Аскорбиновая кислота
Аскорбиновая кислота более известна как витамин С. Аскорбиновая кислота синтезируется из глюкозы в ходе глюкуронового пути. Фермент L-гулонолактон оксидаза, ответственный за превращение гулонолактона в аскорбиновую кислоту, отсутствует у приматов и поэтому аскорбиновая кислота обязательно должна поступать с пищей.
Сам является активной формой. Аскорбиновая кислота участвует в восстановлении различных веществ в самых разных реакциях. Витамин С может восстанавливать цитохромы а и с дыхательной цепи. Самая главная реакция, которая требует витамина С как кофактора, является гидроксилирование пролиновых остатков в коллагене. Следовательно, витамин С требуется для нормального состояния соединительной ткани, а также для заживления рубцов. Т.к. коллаген содержится в органическом матриксе костей, то витамин С также необходим для нормального состояния костей.
Есть и другие реакции, которые требуют витамина С как кофактора. Например, катаболизм тирозина и синтез адреналина из тирозина. Кроме тоо считается, что витамин С связан с процессом стероидогенеза, потому что его болшое количество содержится в коре надпочечников.
Недостаточность в витамине С приводит к болезни, которая называется цинга. Это осуществляется благодаря участия витамина в пост-трансляционной модификации коллагена. Цинга характеризуется мышечной слабостью, кровоточивостью из дёсен, остеропорозом, ломкостью костей, анемией и др. Недостаточность витамина С развивается на фоне либо недостаточного потребления его организмом, либо при повышенной потребности в этом витамине. Повышенное потребление связано с тяжёлым стрессом (или травмой). Это из-за того, что при стрессе происходит истощение коры надпочечников, где много содержится витамина С.
Витамин А
Витамин А состоит из трёх биологически активных молекул: ретинол, ретиналь (ретинальдегид) и ретиноевой кислоты. Каждый из этих трёх компонентов происходит от растительного предшественника бетта-каротина (член семейства каротеноидов). Когда бетта-каротин попал в просвет кишечника, то он расщепляется бетта-каротин диоксидазой и образуется ретиналь. Ретиналь потом восстанавливается до ретинола с помощью ретинальдегид редуктазы. Ретинол этерифицируется пальмитиновой кислотой и вступает в кровь в составе хиломикронов. Потом остатки хиломикронов потребляются печенью, где и происходит запасание витамина А. Транспорт витамина из печени к внепечёночным тканям осуществляется в связанной формой ретинола с апоретинол-связывающим белком. В клетке ретинол связывается с клеточным ретинол-связывающим белком. Для транспорта по крови ретинол связывается с альбумином.
Внутри клеток ретинол связан с рецептором, формируя витамин-рецепторный комплекс. После того, как на рецептор подействует какой-либо сигнал (чаще всего это стероидные и тиреоидные гормоны), этот комплекс взаимодействует с определёнными последовательностями НК в генах, что способствует росту и дифференциации, а также экспрессии этих генов.
Также витамин А участвует в зрении. Световосприятие есть ни что иное, как функция двух типов клеток, расположенных в сетчатке глаза, - это палочки и колбочки. Палочки и колбочки в составе своей мембраны содержат фоторецепторный пигмент. Светочувствительным компонентом в глазах млекопитающих является белок опсин, который ковалентно связан с альдегидом витамина А. Фоторецептором палочек является родопсин, это серпентиновый рецептор. Внутри клеток родопсин связан с G-белком, который здесь называется трансдуцин.
Ретинол также участвует в синтезе определённых гликопротеинов и мукополисахаридов, необходимых для регуляции роста и секреции слизи.
Клинические проявления недостаточности витамина А
Витамин А запасается в печени, и поэтому его недостаточность возникает очень редко. Ранним симптомом недостаточности витамина А является куриная слепота. Потом возникают фолликулярный гиперкератиноз, повышенная чувствительность к инфекциями раку и железо дефицитная анемия. Длительная недостаточность витамина А ведёт к ксерофтальмии (кератинизация роговицы).
Повышенная восприимчивость к раку связана с тем, что бетта-каротин является мощным антиоксидантом.
Витамин Д
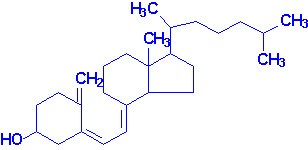
Витамин Д является стероидным гормоном. Он регулирует экспрессию определённых генов. Активной формой гормона является 1,25-дигидроксикальцийтриол. Главной функцией кальцийтриола является регулирование гомеостаза кальция и фосфора. Активный кальцийтриол синтезируется из эргостерола (образуется в растениях) и 7-дегидрохолестерол. Эргокальциферол (витамин Д2) образуется из эргостерола под действием ультрафиолета. В коже 7-дегидрохолестерол превращается в холекальциферол (витамин Д3) также под действием УФ. Холекальциферол абсорбируется в кишечнике и потом транспортируется в печень, связавшись со специфическим витаминД-связывающим белком. В печени холекальциферол гидроксилируется по 25 позиции под действием Д3-25-гидроксилазы и образуется 25-гидрокси-Д3, который является главной формой циркулирования витамина Д. Превращение 25-гидрокси-Д3 в биологически активный кальцийтриол осуществляется с помщью Д3-1-гидроксилазы, который присутствует в проксимальных канальцах почки, в костях и плаценте.
Функция кальцийтриола неразрывно связана с действием паратирина и кальцитонина. Она заключается в регулировании уровня кальция и фосфора. Паратирин освобождается в ответ на низкое содержание кальция и индуцирует продукцию кальцийтриола. Под действием паратирина синтезируется 24.25-дигидроксикальцийтриол. В эпителиоцитах функция кальцийтриола как стероидного гормона заключается в том, что он активирует ген (кальбайндинД28К), который ответственен за синтез белка, который в свою очередь отвечает за абсорбцию кишечного кальция. Увеличение абсорбции кальция требует сопутствующей абсорбции каких-либо отрицательно заряженных ионов для того, чтобы поддерживать электрическое равновесие. Основным таким ионом является Фн. Когда уровень кальция в плазме крови падает, то главным местом для действия кальцийтриола и паратирина является костная ткань, где они стимулируют резорбцию костей, и почки, где они уменьшают выделение кальция посредством стимуляции реабсорбции в дистальных канальцах. Роль кальцитонина в гомеостазе кальция заключается в понижении увеличенного уровня кальция посредством ингибирования резорбции костей.
Клинические проявления недостаточности витамина Д
В тех странах, где витамин Д добавляется в молоко, недостаточность встречается редко. Главными симптомами недостаточности является рахит у детей и остеомалация (размягчение костей) у взрослых. Рахит характеризуется нарушенной минерализацией во время развитие костей, что приводит к размягчению костей. Остеомаляция характеризуется деминерализацией уже сформировавшихся костей, что ведёт к размягчению и повышенной чувсвительности к переломам.
Витамин Е
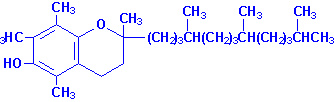
Витамин Е есть ни что иное, как смесь токоферолов. Самым распространённым токоферолом является альфа-токоферол. Витамин Е абсорбируется из кишечника в составе хиломикронов. Доставляется к тканям через транспорт хиломикронов и затем потребляется печенью в составе остатков хиломикронов. Печень может упаковывать витамин в ЛПОНП. Благодаря своей липофильной природе витамин Е накапливается в клеточных мембранах, жировых депо и циркулирующих липопротеинов. Главное место хранения витамина Е - это жировая ткань.
Главная его функция заключается в том, что он является мощным антиоксидантом, связывает свободные радикалы и молекулярный кислород. В частности витамин Е предотвращает от перокисление и чрезмерной десатурации ЖК, которые входят в состав мембран. Альфа-токоферол может связывать два пероксидных свободных радикала и потом связывается с глюкуроновой кислотой и выводится в составе желчи.
Клинические проявления недостаточности витамина Е
Главными симптомами недостаточности витамина Е является увеличение хрупкости эритроцитов, мышечная слабость и бесплодие.
Витамин К
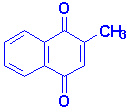
Витамин К существует как К1 (фитилменахинон) в зелёных овощах, как К2 (мультипренилменахинон) в составе кишечной микрофлоре и как К3 в синтетическом менадионе. Главная функция витамина К заключается в поддержании нормального уровня протеинов, которые необходимы для свёртывания крови (факторы 2, 7, 9, 10 и белки С и S). Эти белки синтезируются печенью в неактивном состоянии. Превращение неактивных форм в активные свёртывающие факторы требует посттрансляционной модификации остатков глутаминовой кислоты. Это модификация есть ни что иное, как карбоксилирование остатков глутаминовой кислоты, и фермент, который катализирует эту реакцию требует витамин К как кофактор. В результате образуются гамма-карбоксиглутамат.
В ходе карбоксилирования восстановленная форма витамина К (гидрохинон) превращается в 2,3-эпоксидную форму. Обратная регенерация гидрохинона происходит с помощью редуктазы. Эта последняя реакция является местом действия для дикумарола, который является основой для антикоагулянтов таких как, например, варфарин.
Клинические проявления недостаточности витамина К
Витамин К всасывается в кишечнике только в присутствии желчных солей. Частично витамин К синтезируется микрофлорой кишечника, поэтому все те лекарства, которые действуют на микрофлору, могут привести к недостаточности. Основным симптомом является геморрагический синдром
Гормоны
Гормоны - класс регуляторных химических соединений, синтезируемых клеткой. Термин hormone ввёл в 1905 году Старлинг что означает возбуждать, но есть и тормозные гормоны, которые угнетают какую-либо функцию.
Принцип секреции гормона:
-эндокринная секреция - гормон попадает в кровь;
-паракринная - гормон не попадает в кровь, а действует на соседние клетки;
-аутокринная - клетка выделяет гормон для себя;
-нейроэндокринная - нервная клетка выделяет не только медиатор, но и гормон, который идёт в кровь.
Гормоны классифицируют по их химической природе:
-белки и пептиды (инсулин, глюкагон и др.);
-производные аминокислот (Т3, Т4, адреналин, НА - из тирозина, серотонин - из триптофана, гистамин из гистидина);
-стероиды - являются производными холестерина.
Классификация рецепторов
Рецепторы делят на три класса:
1).Рецепторы, которые пронизывают мембрану и обладают ферментативной активностью. Эти рецепторы обладают активностью одного из ферментов:
-тирозин киназа (инсулин);
-тирозин фосфатаза (СD45);
-гуанилат циклаза (NO);
-серин/треонин киназа.
Рецепторы с тирозинкиназной активностью способны к автофосфорилированию точно также, как и другие субстраты.
Кроме того, есть рецепторы, которые сами по себе не обладают ферментативной активностью, но связаны с внутриклеточной тирозин киназой посредством прямого белок-белок взаимодействия.
2).Рецепторы, которые внутри клеток связаны с белком, который в свою очередь связан с ГТФ (поэтому этот белок и называется G-белком). Эти рецепторы семь раз пронизывают мембрану и называются серпентиновыми (от англ. serpent - змея). Сюда относятся адренергические рецепторы, рецепторы к одорантам и большинства гормонам (глюкагон, ангиотензин, вазопрессин и др.)
3).Рецепторы, которые расположены внутри клеток. Как только лиганд взаимодействует с таким рецептором, то формируется лиганд-рецепторный комплекс, который мигрирует в ядро, где напрямую действует на транскрипцию генов. Сюда относятся рецепторы к стероидным гормонам и тиреоидным. Эти гормоны могут свободно проникать внутрь клетки.
Молекулярный механизм действия
Рецепторы ко всем белковым гормонам расположены на поверхности клеток. Как только гормон присоединяется к рецептору, то последний изменяет свою конформацию и взаимодействует с G-белком. При этом ГДФ на альфа-субъединице G-белка заменяется на ГТФ. Затем G-белком отсоединяется от гормон-рецепторного комплекса и диссоциирует на альфа- и бетта,гамма-субъединицы. Gа-ГТФ связывается с эффекторным белком (ЭБ). ЭБ - это фермент, который катализирует реакцию образования вторичнюх посредников. Эффекторным белком может быть аденилатциклаза, фосфолипаза и др.
А. При ЭБ - аденилатциклазе. Этот фермент катализирует образование цАМФ. цАМФ активирует протеинкиназу А, которая регулирует фосфорилирование множества ферментов.
Б. ЭБ - фосфолипаза С. Этот фермент катализирует реакцию образования ИФ3 и ДАГ из фосфатидилинозитол-4,5-дифосфата (ФИДФ). Затем ИФ3 связывается с ЭПР, открывая кальциевые каналы, что ведёт к транспортировке кальция в цитоплазму. Ионы кальция могут оказывать прямое действие, активируя некоторые ферменты, либо опосредованное, через специальные белки, связывающие с кальцием. Один из таких белков - кальмодулин. ДАГ выполняет две функции: активирует протеинкиназу С и открывает кальциевые каналы, усиливая эффект ИФ3. Как и ПКА, протеинкиназа С фосфорилирует треониновые и сериновые остатки многих белков.
Дата добавления: 2016-02-24; просмотров: 1818;