Уровня СМФП в области левой (s.) и правой (d.) миндалин (Amygd.) в период 5 страница
Декодирование сигналов означает восстановление начального сообщения.
3. Под кодом понимают в частном случае определенную систему символов.
4. Выбор наиболее экономичного кода обеспечивает быструю передачу сообщения, информации, однако при этом вероятность искажения сигнала увеличивается. Чтобы уменьшить эту вероятность, передают избыточную информацию.
5. Кодированием называется процесс преобразования одного сигнала в другой, наиболее пригодный для передачи информации по конкретному каналу связи, - процесс установления определенного соответствия между сообщением и той системой символов, при помощи которых эти сообщения передаются.
6. В более широком смысле под кодированием понимается любое сопоставление, в том числе одних знаков или сигналов с другими.
Этими формулировками понятие «кодирование» не исчерпывается. Тех, кто дополнительно интересуется именно терминологической стороной проблемы, отсылаем к книге Сомьена (Somjen, 1975).
К какому классу процессов целесообразнее всего отнести паттерны, описанные в наших статьях, монографии и в данной книге? Они безусловно подходят под всеобъемлющее понятие «коррелят», однако именно в данном случае это понятие в связи с тем, что обычно вкладывается в указанный термин, не отражает уровня изучения вопроса. Паттерн, выделенный из импульсной активности, обладает той степенью характерности, которая позволяет использовать его в качестве эталона при эталонном машинном поиске аналогов этого паттерна в импульсной активности. Нейрофизиологические корреляты, как правило, менее специфичны и, отражая в нервной системе какое-то конкретное явление, за очень редким исключением, по существу, отражают класс явлений. Так, например, местные медленные волны на ЭЭГ указывают на изменения состояния нервной ткани, развивающиеся при опухолях мозга. Но если заложить в качестве эталона в ЭВМ отрезок записи медленноволновой активности, то окажется, что машина выловит из ЭЭГ сообразно с эталоном медленные волны и при опухолях, и при очаговых сосудистых поражениях, и при травме, и при воспалительном процессе, и во время сна здорового человека. Более специфичным окажется в качестве эталона наблюдающийся при эпилепсии комплекс острой и медленной волн (спайк-волна). В этом случае машина быстро обнаружит его
аналоги, и без того хорошо видные на ЭЭГ невооруженным глазом.
Однако в случае тех паттернов, о которых идет речь, заложив один из них в ЭВМ, на отрезке записи от десятков секунд до десятков минут можно опознать момент появления именно его аналога в импульсной активности, например, заложив эталон, соотносимый с другим словом, импульсную перестройку, сопоставимую с этим другим словом, и т. д. Паттерны оказываются приемлемыми для использования в качестве эталонов при машинном поиске, они различны для разных групп слов, и, таким образом, если рассматривать эти паттерны как корреляты слов, приходится признать, что это корреляты более высокого, чем обычно, порядка. Но, как показано выше, паттерн, выделенный из импульсной активности, может быть расщеплен на составляющие части, по крайней мере, часть из которых обладает достаточной характерностью для использования в качестве эталона при машинном поиске аналогичных паттернов в импульсной активности нейронных популяций. Таким образом, паттерн все же достаточно характерен, как и некоторые его элементы. При использовании такого элемента в качестве эталона в импульсной активности обнаруживаются те паттерны, которые имеют в своем составе данный элемент и соотносятся с отражением в мозгу определенных слов. Используя этот эталон при анализе фоновых записей, естественно, выявляли единичные аналоги его, но анализ записей импульсной активности, осуществленных при восприятии, удержании в памяти, воспроизведении и обобщении других слов, с помощью данного эталона был малоэффективен.
Рассматривая данные анализа мультиклеточной активности, следует разобраться, почему именно триада рассматривается как единица паттерна. Действительно, в импульсной активности выделяются и другие регулярные элементы - повторяющиеся последовательности с меньшим или большим количеством определенных интервалов. Так, например, в импульсной активности парных интервалов наблюдается значительно больше, чем триад.
При одновременной обработке достаточно большого количества нейронов из активной популяции оказывается, что и в фоне (редко), и в момент теста обнаруживаются воспроизводимые групповые последовательности разрядов, но уже не с тремя, а с большим числом определенных интервалов. И все же есть ли достаточно убедительные основания для того, чтобы считать именно триаду основной или единственной элементарной единицей паттерна? Единственной - нет.
Паттерн может содержать и связанные пары интервалов, и большие регулярные группы разрядов. Элементарной единицей паттерна может быть и последовательность из четырех и более интервалов. Что, однако, важно подчеркнуть при рассмотрении данного вопроса - именно триада может быть минимально достаточной характерной единицей паттерна. Именно триады являются и более устойчивыми элементарными единицами, хотя это не исключает значения элементов с большим количеством последовательных разрядов.
В паттерне, соотносимом со словом или его элементом, как указывалось в ряде проведенных нами исследований, может быть обнаружено множество триад. Проверить степень специфичности каждой из них, время устойчивости первоначально возникшего множества элементов паттерна нелегко, но возможно с помощью уже имеющихся методов. В работах Н. Г. Шкуриной, исследовавшей
нейрофизиологическое отражение синтетических фонем, показано, что и этот полный комплекс - паттерн - может быть использован для дифференцировки отражения разных фонем. Следует помнить, что отдельные триады при эталонном поиске, наоборот, могли быть использованы для выделения группы слов общего смыслового поля. Отсюда, если целесообразна проверка значимости и характерности отдельных элементов, то целесообразна и своего рода обратная операция - исследование динамики характерности паттерна при последовательном изменении количества триад, вводимых в паттерн и выводимых из него. Именно на этих путях лежит решение вопроса о том, какой набор элементов в одной нейронной популяции является необходимым и достаточным отражением одного слова. Однако, зная хотя бы некоторые из общих механизмов мозга, следует учитывать, что избыточность не означает ненужности.
Далее, как указывалось выше, характерные перестройки импульсной активности - паттерны - наблюдаются в исследуемых условиях не в одной, а в нескольких популяциях, в нескольких зонах мозга, а если учесть условия исследования, повидимому, правомерно сказать - во многих зонах. Как соотносятся паттерны разных зон мозга?
Организация импульсной активности в одних и тех же условиях варьирует в разных зонах мозга и в так называемой фоновой записи - до предъявления теста, и при восприятии слов, словоподобных сигналов, слогов и фонем. Выделение из общего паттерна его элементарных составляющих показывает, что большая часть их оказывается различной в разных зонах мозга. Однако имеются и сходные по последовательным межимпульсным интервалам группы. Рабочей гипотезой (Бехтерева, 1977), объясняющей на сегодня эти факты, является представление о распределенности в мозгу полного паттерна - отражения - ио значении дублирующих элементов как системообразующих, организующих систему факторов. Эта гипотеза может проверяться прежде всего путем исследования функционального значения составляющих паттерна методом эталонного поиска. Однако и уже имеющиеся материалы по динамике паттерна в процессе более сложной психической деятельности заставляют предположить, что дело, особенно в отношении синхронных элементов в разных зонах мозга, обстоит если не иначе (для такого мнения нет оснований), то сложнее.
Таким образом, в нашем случае использование терминов «код» и «кодирование» основано прежде всего на характерности выделяемых короткоживущих паттернов или по крайней мере некоторых их элементов, позволяющей на определенных отрезках времени не только наблюдать появление этой перестройки в мозгу при одном и том же внешнем сигнале (слове и т. д.), но и определять в импульсной активности моменты возникновения соответствующих отражений слов, словоподобных сигналов и их элементов, используя предварительно выделенный их аналог в виде эталона при основанном на сравнении с этим эталоном машинном поиске. Фактами, на которых основывается представление о кодовом значении регистрируемых паттернов, является и функциональная значимость их элементов, и возможность встраивания элементов в паттерн в специальных условиях исследования. По-видимому, и сама динамика паттерна по ходу простейших тестов на восприятие и воспроизведение слов свидетельствует также о кодовом значении этих паттернов: паттерн возникает в момент восприятия слова, далее претерпевает различные трансформации в зависимости от базиса долгосрочной памяти и вновь появляется перед произнесением и во время произнесения слов. Кодовое значение паттернов подчеркивается наличием в паттернах разных зон мозга сходных элементов. И вто же самое время использование в данном случае «кодовой» терминологии требует во избежание недоразумений пояснений и постановки ряда вопросов, решение которых важно для дальнейшего изучения проблемы мозгового кода психической деятельности.
Так, и это, пожалуй, самое основное - исследование активности многих нейронных популяций подчеркнуло, что регистрируемые паттерны - это местные явления в мозгу. Во всяком случае паттерн в таком виде не передается с соответствующих анализаторных, в данном случае слуховых, образований мозга, а возникает местно под влиянием факторов, рассмотренных выше. В таком полном виде, по-видимому, паттерн не передается и дальше. Это положение является не только результатом логического сопоставления массивности и сложности данного элемента с передаточными возможностями каналов связи и временными параметрами очень быстрых процессов - психических явлений, но и тех же исследований паттернов в большом числе разных нейронных популяций мозга. Обнаруженные и исследованные нами паттерны - местные, привязанные к структуре тончайшие нейрофизиологические корреляты психической деятельности или, точнее, нейрофизиологические выражения кодирования и декодирования, детерминированные в значительной мере памятью и обучением. Это - та нейродинамика (или ее часть), возникновение которой находится в прямой зависимости от участия структуры в обеспечении психической деятельности. Естественно, очень важно знать, как, на основе каких процессов осуществляется взаимодействие между структурами в высших отделах нервной системы? Но это уже другой вопрос, хотя и связанный с проблемой местного кода. Он будет рассмотрен нами далее.
Пока еще трудно с уверенностью сказать, какой именно уровень интеграции обеспечивается деятельностью исследуемых структур стриопаллидарной системы, зрительного бугра, верхних отделов ретикулярной формации (весьма, кстати, дифференцированной - П. Г. Костюк, 1973), медиобазальных отделов височной доли, премоторной и некоторых других отделов коры. Как будто трудно отрицать на основе все увеличивающегося числа экспериментальных функционально-анатомических данных и исследований, проводимых у человека в разных лабораториях мира, что изучаемые подкорковые зоны мозга значительно более специфичны, чем это предполагалось ранее, и если не являются единственными областями высшей интеграции при осуществлении психической деятельности, то, безусловно, в ней очень активно участвуют (Костандов, 1978, и др.). В рецепторных и анализаторных образованиях идет первичное кодирование сигнала, перекодирование, адаптированное к передаче, перекодирование для первичного анализа. Этот вопрос подробно рассматривается в работах Уордена и Марша (Worden, Marsh, 1968), Я. А. Альтмана (1972), Кейдела (Keidel, 1974), Моллера (Moller, 1974, 1978а, 19786), Михольсонаи Винсента (Michelson, Vincent, 1975), Десмедта (Desmedt, 1977), Мушегиана с соавторами (Moushegian et al., 1978), Л. А. Чистович (1978) иее сотрудников и многих других. В высших по отношению к анализаторным структурах, в которые стекается информация от разных анализаторов по прямым и непрямым путям, происходит или, точнее, может происходить расшифровка свойств более сложных сигналов, в данный момент или онтогенетически воспринимавшихся через разные анализаторные входы.
В них должен осуществляться уже не только процесс кодирования, но и обязательно - местного декодирования сигнала, формирования местных, в пространстве мозга взаимодополняющих оперативных единиц интегративной деятельности. Этот декодированный сигнал уже не ограничен необходимостью обладать свойствами, приемлемыми для передачи по нервным путям. Его взаимодействие с аналогичным сигналом в остальных задействованных зонах мозга идет по другим законам, а поэтому он может иметь и существенную избыточность. Декодированный сигнал, с одной стороны, изменяет состояние структуры и, с другой - проявляется только на определенном ее функциональном фоне, выявляет свойства структуры, считывающей и шифрующей его в биохимическом коде и вновь «отдающей» его в биоэлектрический код - по требованию.
Паттерны-коды и их элементы - групповые последовательности разрядов - удерживаются или воспроизводятся в том же виде в нейронной популяции на протяжении многих секунд и даже десятков минут в зависимости от характера теста, свойств структуры и ее функционального состояния. Период жизни этих нейродинамических единств, таким образом, ограничен. В первую очередь это относится к групповым последовательностям разрядов с определенными интервалами. Далее может развиться реорганизация импульс ной активности данной нейронной популяции, местная или отражающая общие перестройки в мозгу, и при сохранении того же принципа в паттерне, соответствующем тому же слову, будут возникать и удерживаться на некоторое время те же или уже другие групповые последовательности разрядов, на данный новый отрезок времени также с определенными интервалами. Паттерны и их элементы формируются, удерживаются и распадаются с сохранением в фоновой активности опорных элементов. Популяция нейронов оказывается вновь готовой к деятельности, формированию по тем же принципам новых паттернов и их элементов. Принципиальная возможность этого подчеркивается наличием в мозгу человека нейронов, дифференцированно реагирующих на семантику вербальных тестов (Гоголицын, 1979; Кропотов, 19796) (рис. 17). В исследовательском плане данные этих авторов определяют поиск мозговых зон с более и менее устойчивыми элементами паттерна и одновременное использование элементов и полного дистрибутивного паттерна при изучении нейрофизиологической организации восприятия, удержания и воспроизведения слов и осуществления мыслительных процессов (Шкурина, 1983).
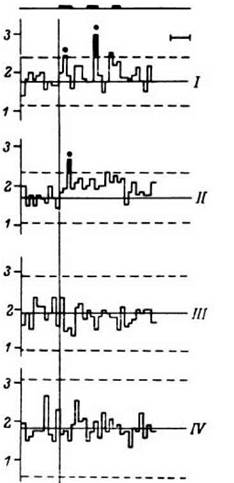
Рис. 17. ПСГ активности одиночного нейрона крючка извилины гиппокампа. Каждая гистограмма накоплена по 12 предъявлениям пар слов, которые обобщались по смыслу пациентом. III - гистограмма, накопленная при предъявлении цифры. Остальные обозначения - как на рис. 16. Видно отсутствие ответа на гистограммах III и IV, а также некоторые различия в положении максимумов на гистограммах I и II (черные кружки)
Подчеркивание динамичности биоэлектрических коррелятов психической деятельности, их неразрывной связи со свойствами мозговых структур, в которых они регистрируются, важно для понимания того, что дальнейшее изучение мозгового кода психических процессов требует не только одновременного использования ряда составляющих комплексного метода изучения мозга, но и прежде всего сопоставления импульсной активности нейронных популяций с медленными электрическими процессами. Оно требует также обязательно сочетанных физиологических и молекулярно-биохимических исследований. Именно в последние годы и этот комплекс становится все более реальным.
На основе использования главным образом показателя структуры импульсного потока были проведены и другие исследования, целью которых являлось получение хотя бы ориентировочного представления о нейродинамике при реализации несколько более сложных психологических тестов. Подробно эти данные приведены нами в первом издании данной книги (1980).
Суть этих работ в изучении способом эталонного поиска того, что развивается в динамике импульсной активности нейронных популяций в условиях, когда испытуемым предъявлялись цветные пейзажи или жанровые картинки в виде слайдов и предлагалось ответить, например, на вопрос о том, к какому сезону года относится данный снимок, или на аналогичный вопрос, детерминирующий принятие решения. Исследование строилось таким образом, что дистрибутивные паттерны - корреляты слов, служащие в дальнейшем эталоном для машинного поиска, выделялись во время однократного исследования продолжительностью в несколько минут и реже - десятков минут с предъявлением этих слов и в том же исследовании реализовалась задача с указанным выше принятием решения. Такие жесткие условия были необходимыми в связи с обнаруженной и описанной динамичностью дистрибутивных паттернов и их элементарных единиц - интервальных последовательностей. Полученные нейрофизиологические данные принципиально не отличались от того, что было приведено ранее в связи с исследованиями, где предъявлялись квазислова, слова, слоги, фонемы и тесты на обобщение слов, имеющих смысловую общность. В импульсной активности нейронных популяций исследованных нами подкорковых структур при рассмотрении слайда и принятия по нему задаваемого решения при эталонном машинном поиске обнаруживались перестройки структуры импульсного потока, выявленные ранее (в том же тесте!) при прослушивании и/или произнесении больным слов, имеющих отношение к содержанию слайда или ответу.
Большинство такого рода работ было проведено нами в 1970-х годах, когда важнейшим вопросом была принципиальная проходимость или, наоборот, непроходимость пути.
В результате этих исследований были получены первые опорные данные о возможности извлечения из динамики импульсной активности коррелятов
мыслительных процессов. Большим достоинством этих исследований, которые было бы неправомерно недооценивать, явилось изучение того, что происходит в мозгу в ходе единичного опознания сигнала, однократного принятия решения и т. д., то есть соответствие физиологического исследования тому, как решает свои проблемы мозг человека. В то же самое время динамичность исследованных перестроек импульсной активности, возможность ориентации на них лишь в коротких временных промежутках и, таким образом, полное отсутствие рутинно требуемой статистической подтвержденности определили следующий этап работы.
Второй этап исследований. Суть этого, второго, этапа наших работ в той же проблеме изучения нейрофизиологических коррелятов мыслительной деятельности была в решении задачи получения максимально достоверных данных о мозговой нейродинамике при мыслительной деятельности. Это отражало развитие работ, оно было важно для того, чтобы данные обрели необходимую устойчивость и могли быть воспроизведены в любой другой, располагающей соответствующими возможностями лаборатории. Своего рода упрощение в этих работах, однако, определялось тем, что при анализе импульсной активности как бы отступали от того, как реализуется мыслительная деятельность взрослого человека, отступали от изучения нейродинамики, коррелирующей с одиночными тестами. Исследовался так называемый накопленный ответ мозга при предъявлении достаточно большого числа (до 60 и даже более 100) принципиально однотипных проб, соотносимый с фоновой импульсной активностью, сравниваемый в соответствии с фазами теста, его вариациями и характером выполнения проб. Короче говоря, множество различных вопросов решалось при изучении местных процессов способом перистимульных гистограмм (ПСГ). Следует, однако, подчеркнуть, что одновременно, хотя и менее интенсивно, изучалась структура импульсного потока и весьма интенсивно - организация системы обеспечения мыслительной деятельности.
Таким образом, на данном этапе работы основная задача осталась той же - исследование отражения мыслительной деятельности в перестройках импульсной активности нейронных популяций коры и подкорки. Одним из важных отличий от первого этапа была существенная стандартизация тестов, необходимая при исследовании динамики импульсной активности методом ПСГ, обязательным условием которого являлось накопление данных. Испытуемым предъявлялись пробы в виде слайдов в зависимости от задачи с помощью тахистоскопа, светодиодной матрицы или в виде звуковых сигналов. В задачах на опознание в случайном порядке предъявлялись изображения (точнее, силуэты), имеющие смысловое значение, и изображения, одинаковые с первыми по физическим характеристикам, но не имеющие смыслового значения (рис. 18). Изображения, в том числе и цифры, предъявлялись с достаточным для опознания временем или таким образом (по времени), что правильное опознание регистрировалось примерно в 50 % проб.
'И’йТ
1?Rtf
f тимуА
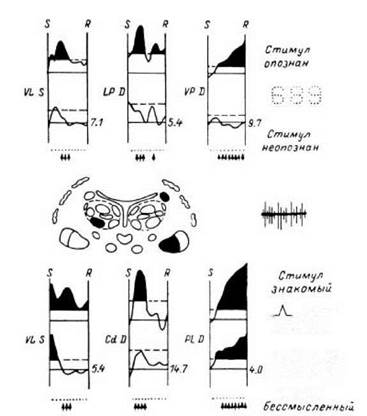
рис Примеры пар ПСГ, вычисленных для нейронных популяций в различных образованиях таламуса и стриопаллидарной системы, в пробах с предъявлением цифр на пороге опознания (вверху) и в пробах с предъявлением знакомых и бессмысленных многоугольников (внизу).
S - предъявление стимула; R - начало вербальной реакции больного. VL S - вентролатеральное ядро левого полушария; LP D, VP D, Cd D, Pl D - соответственно заднее латеральное, заднее вентральное, хвостатое и подушечное ядра правого полушария. Точки - отметки времени (бин - 150 мс). Цифры справа внизу каждой ПСГ - средняя частота импульсов в бине. Стрелки указывают на бины, в которых обнаружены значимые различия между средними частотами разрядов для верхней и нижней гистограмм. Непрерывные горизонтальные линии - средние уровни частоты разрядов в фоновом фрагменте, прерывистые- уровни значимости Р < 0.01 отличия от частоты в фоновом фрагменте. Фрагменты гистограммы, выходящие за пределы уровней значимости, отмечены черным цветом
В нейронных популяциях, расположенных в области VA, VL, LP, Cd, CC, Ci, на ПСГ наблюдались коротколатентные компоненты (латентный период до 200 мс), одинаковые при предъявлении и значимых, и незначимых силуэтов изображений. В области VA, VL, LP, Rt, CM, Cd, CC, Ci наблюдались один или два типа длиннолатентных компонентов, причем первый из них (латентный период свыше 250 мс), так же как и коротколатентный, имел форму, близкую к колоколообразной, а второй мог быть характеризован как медленное учащение (или урежение) разрядов
нейронов. Эти длиннолатентные компоненты наблюдались при предъявлении значимых стимулов (см. рис. 14). В задачу обследуемого лица в данном тесте входило опознание сигнала, дача ответа, соответствующего изображению в случае значимого сигнала, и ответа «нет», если значение сигнала было неизвестным. Медленное учащение разрядов наблюдалось непосредственно перед словесным ответом и, возможно, характеризовало подготовку к нему.
Таким образом, было обнаружено длиннолатентное учащение разрядов нейронов различных зон мозга, статистически достоверно связанное с опознанием значимых изображений, - отражение в реакции нейронов смыслового элемента проб (Кропотов в кн.: Бехтерева и др., 1985). Для дальнейшего изучения именно этой важнейшей в наших работах позиции сотрудником лаборатории Ю. Д. Кропотовым (1984) проводилось специальное исследование, в котором бессмысленным изображениям- силуэтам придавался определенный смысл. Больной обучался узнавать его как имеющий определенное значение. В импульсной активности нейронных популяций, расположенных в области вентролатерального ядра таламуса, на предъявление ранее незначимого сигнала появились длиннолатентные компоненты (рис. 19). Эти наблюдения еще раз подтвердили, что длиннолатентные компоненты ПГС связаны с отражением смысловых элементов реализуемых проб.
Мне казалось целесообразным здесь достаточно подробно остановиться на этих исследованиях, как существенно подтверждающих своего рода «хрупкие» данные первого этапа. Ивто же самое время, как вполне понятно, данная книга не является эпическим повествованием обо всем том действительно огромном материале, который накоплен за годы исследований нейрофизиологии мыслительной деятельности в лаборатории.
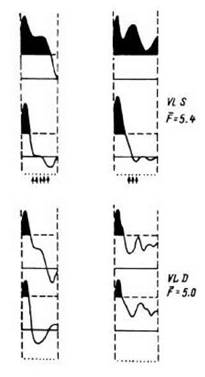
Рис. 19. Сравнительный анализ ПСГ для двух
нейронных популяций больного паркинсонизмом, выполнявшего тесты на опознание и обучение.
Слева - пары ПСГ для теста на обучение (верхние гистограммы в каждой паре - суммарный вызванный ответ на первые четыре предъявления стимула, нижние - вызванные ответы на последние четыре предъявления). Справа - пары ПСГ для теста на опознание (верхние гистограммы, в каждой паре - предъявление семантически значимых, нижние - семантически незначимых стимулов). Остальные обозначения - как на рис. 18
Поэтому, например, в отношении исследований с предъявлением сигналов на пороге опознания будут приведены только некоторые данные, а в случае, если читатель захочет подробнее ознакомиться именно с этим вопросом, он может обратиться непосредственно к работам сотрудников нашей лаборатории (Кропотов, 1983а, 19836; Бехтереваи др., 1985а).
Пробы с предъявлением сигналов на пороге опознания были сконструированы таким образом, что, как указывалось, в 50 % случаях больной не опознавал сигнал или опознавал его ошибочно. В данном случае в прямой зависимости от исследуемой зоны мозга наблюдалась одинаковая активационная (учащение разрядов) или тормозная (урежение разрядов) реакция или избирательная реакция: только на правильное и, что очень важно подчеркнуть, только на неопознание или ошибочное опознание сигнала (рис. 20).
Сопоставимые с этими наблюдениями данные были получены нами впервые вместе с В. Б. Гречиным в 1968 году. При регистрации медленных физиологических процессов мозга - таких, как локальный кровоток и напряжение кислорода, осуществляемой во время выполнения психологических тестов, было обнаружено, что в некоторых зонах хвостатого ядра и таламических образований изменение этих процессов обнаруживалось избирательно при ошибочной реализации теста.
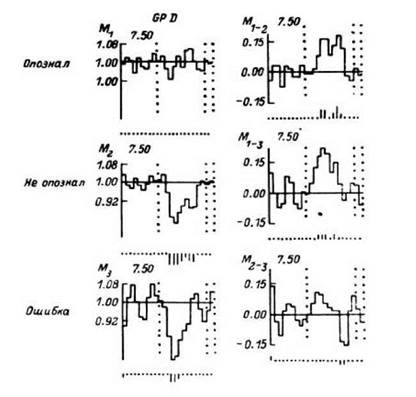
Рис. 20. ПСГ импульсной активности нейронной популяции бледного шара правого полушария для случаев опознания (М1), неопознания (М2) и ошибочного опознания (М3) зрительных стимулов (цифр), предъявляемых на пороге опознания, а также различия между ПСГ (М1-2, М1-3, М2-3).
На M -по оси ординат - отношение средней частоты разрядов данного бина На M1—М3 по оси ординат - отношение средней частоты разрядов данного бина к средней частоте фонового фрагмента. На М1-2, М1-3, М2-3 по оси ординат - отношение разности средних частот разрядов соответствующих ПСГ к средней частоте разрядов в данном бине. Под гистограммами - отметки бинов с символическим отражением уровней значимости в длине вертикальных отрезков (Р < 0.05; Р < 0.01; Р < 0.001). Левый вертикальный пунктир - предъявление цифры, два вертикальных пунктира справа - предъявление триггерного стимула, служащего сигналом для вербальной реакции испытуемого. Цифры над гистограммами - среднее число импульсов в бине для фонового фрагмента
Это явление было названо нами детекцией ошибок и соответственно зоны, где оно обнаруживалось, - детекторами ошибок. Конструкция психологических тестов в тех ранних исследованиях не предполагала большого количества ошибок, они были редки
- и соответственно еще реже наблюдалась детекция ошибок. Феномен оставался описанным, но трудно было на основании исследования медленных процессов судить о физиологической сущности эффекта. При анализе импульсной активности нейронов в условиях реализации тестов указанной выше конструкции феномен стал «ручным», его можно было наблюдать достаточно большое количество раз в пределах теста для того, чтобы получить статистически достоверные результаты. В зонах мозга, расположенных в области бледного шара, неопознание и ошибочное опознание сигналов соотносилось с фазическим урежением импульсной активности - с реакцией тормозного типа. Следует отметить, что реакции тормозного типа наблюдались в ряде областей мозга и, в частности, в том же бледном шаре и при правильных реализациях теста, и менее селективно - вне зависимости от характера опознания.
Ранее (Бехтерева, Гречин, 1968; Бехтерева, 1971) нами предполагалось, что детекция ошибок является оптимизирующим механизмом мозга, способствующим облегчению последующих мыслительных операций. Местная тормозная реакция в областях мозга, где менее селективные реакции тормозного типа также наблюдаются, может рассматриваться также как элемент оптимизирующих механизмов мозга, хотя вопрос, несомненно, нуждается в дальнейшем уточнении и исследовании. Косвенным свидетельством того, что в данном случае есть некоторые основания думать о торможении тормозных зон, являются наши наблюдения, впервые опубликованные в 1964 году и позднее подтвержденные, о том, что электрическая стимуляция некоторых зон хвостатого ядра и других областей мозга вызывала клинические и поведенческие тормозные реакции.
Вопрос о физиологическом значении явления детекции ошибок интересен в плане изучения общих и частных механизмов мозга и, несомненно, заслуживает дальнейшего исследования. Здесь возможно и важно проследить влияние различных фармакологических средств, а также уточнить, является ли данный феномен специальным атрибутом системы обеспечения мыслительной деятельности (что маловероятно) или это более общий механизм мозга. Заканчивая на сегодня разговор о детекции ошибок, пожалуй, можно сказать, что, каково бы ни было значение этого явления, селективность реакций импульсной активности при психологических тестах в зависимости от качества их выполнения косвенно также свидетельствует в пользу зависимости динамики импульсной активности от содержательных (смысловых) элементов психических процессов. Статистически достоверные данные об изменениях импульсной активности в связи со смысловыми характеристиками теста были получены способом ПСГ и при пробах, требующих реализации относительно более сложных мыслительных операций. Естественно, как и большинство психологических проб, удовлетворяющих условиям многократного повторения в тесте, они были сложнее действительно лишь относительно. Абсолютно они были очень просты, однако, что важно, требовали постоянного внимания и хотя и элементарного, но все же принятия решения о характере ответа.
Дата добавления: 2016-02-20; просмотров: 586;