Деятельность человека и
Эволюция биосферы
Э. И. Колчинский (1988) в эволюции биосферы выделяет следующие тенденции: постепенное увеличение общей ее биомассы и продуктивности; прогрессивное накопление аккумулированной солнечной энергии в поверхностных оболочках Земли; увеличение информационной емкости биосферы, проявляющейся в нарастающем росте органических форм, увеличении числа геохимических барьеров и возрастании дифференцированности физико-географической структуры биосферы; усиление некоторых биогеохимических функций живого вещества и появление новых функций; усиление преобразующего воздействия жизни на атмосферу, гидросферу, литосферу и увеличение роли живого вещества, продуктов его жизнедеятельности в геологических, геохимических и физико-географических процессах; расширение сферы действия биологического (биотического) круговорота и усложнение его структуры. Несомненно, к этому перечню необходимо отнести трансформирующее воздействие на биосферу человеческой деятельности, не исключая нисходящую ветвь эволюции биосферы — все эволюционирующие системы не являются бессмертными, а имеют «начало» и «конец» своего существования. Так, если в эволюции живого вещества имеется непрерывный поток генетической информации, а в геноме человека есть гены от всего ряда его предков, то в составе биосферы имеются виды различного географического возраста — «эиогеноэлементы», или «биоэлементы», экосистем. Происходит эволюционная замена данных экогеноэ-лементов (биоэлементов), иногда в региональных рамках полная замена, с исчезновением предшественников.
Не могло не изменить естественных процессов массовое истребление человеком растений и животных, к примеру, плиоценовое исчезновение крупных животных, по всей вероятности, происходило не только из-за прямого преследования, но и в результате нарушения цепей питания, в целом пищевых сетей, что вело к преобразованию экосистем. Современное уничтожение видов, которое идет намного быстрее, чем во времена плиоценового перепромысла, должно вести и ведет к процессам, обратным к названным Э. И. Колчинским — снижается биомасса, продуктивность и информационность биосферы, меняется характер аккумуляции солнечной энергии в поверхностных оболочках планеты и т. п. Отсюда закономерности эволюции биосферы необходимо рассматривать как в прогрессивном, так и в регрессивном плане.
Как нам уже известно, эволюция живого началась с возникновения форм преджизни, а в дальнейшем и праорганизмов (рис. 12.44).
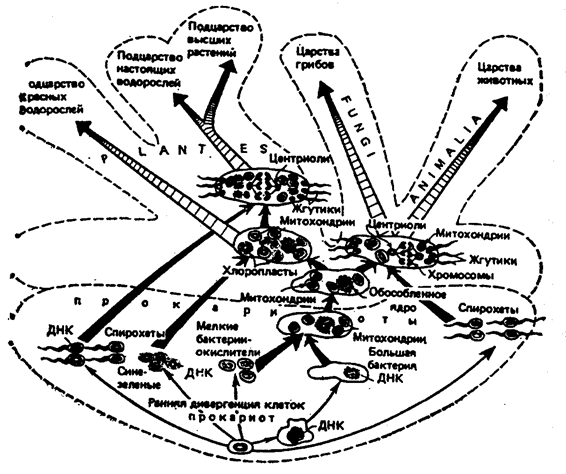
Рис. 12.44. Последовательность этапов симбиогенетического
происхождения клеток эукариот, наложенная на
родословное древо клеточных (кариот)
Примечание: показана неоднократность этапов симбиогенеза клетки эукариот
С этого геологического времени начал действовать принцип Реди: живое происходит только от живого, между живым и неживым веществом существует непроходимая граница, несмотря на то, что имеется постоянное взаи-модействие. В дальнейшем данное обобщение было заново сформулировано В. И. Вернадским в 1924 г. Именно этот принцип служит подоснов Д вой сложения экосистем в рамках таких закономерностей, как разграничение между живым и неживым. Взаимосвязь между ними формирует дополнительность и соответствие внутри биотического сообщества и связь биотоп — биоценоз.
В реальной эволюции принцип Реди проявляется весьма многообразно—способы видообразования, сложения био-, экобио- и экосистем многочисленны, хотя и подчиняются общим законам биологической микро- и макроэволюции, а также экогенез. На разных этапах развития биосферы процессы в ней не были одинаковыми, несмотря на то что шли по аналогичным схемам. Наличие ярко выраженного круговорота веществ, согласно закону глобального замыкания биогеохимического круговорота, является обязательным свойством биосферы любого этапа ее развития. Вероятно, это непреложный закон ее существования. Следует особо обратить внимание т увеличение доли биологического, а не геохимического, компонента в замыкании биогеохимического круговорота веществ. Ныне существующий тип биогеохимического обмена, состоящий из автотрофов-процудентов, гете-ротрофов-консументов и гетеротрофов-редуцентов со все большим ростом управляющего значения среднего звена, практически сложился в середине мелового периода. Если на первых этапах эволюции преобладал общебиосферный цикл — большой биосферный круг обмена (сначала только в пределах водной среды, а затем разделенный на два подцикла — суши и океана), то в дальнейшем он стал дробиться. Вместо относительно гомогенной биоты появились и все глубже дифференцировались экосистемы различного уровня иерархии и географической дислокации. Приобрели важное значение малые, биогеоценотические, обменные круги. Возник так называемый «обмен обменов» — стройная система биогеохимических круговоротов с высочайшим значением биотической составляющей.
Деятельность человека ведет к гомогенизации систем биосферы. Все больше «стираются» элементарные экосистемы, превращаясь в «монотонные» агросистемы, однообразные по биогеохимическим характеристикам культурные ландшафты. При этом снижается степень замкнутости биогеохимических циклов. Вероятно, в этом заключается секрет накопления в биосфере, и в первую очередь в атмосфере, малых газовых примесей, выброса тех веществ, которые, естественно, образуются в меньшем количестве и обычно ранее утилизировались биотой практически полностью. Чем больше организмы воздействовали на среду биосферы, тем интенсивнее шла эволюция. Этот принцип максимума эффекта внешней работы, закон саморазвития биосистем или закон исторического развития биологических систем, был сформулирован в 1935 г. Э. Бауэром: развитие биологических систем является результатом увеличения их внешней работы — воздействия этих систем на окружающую среду.
Физико-математическое подтверждение вышеназванных обобщений дает теорема порога возрастания энтропии в биосфере, или теорема К. С. Тринчера, выведенная в 1964 г., — продукция энтропии живым веществом биосферы возрастает до порога, определяемого уравнением:
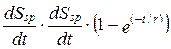 , (12.9)
, (12.9)
где t — абсолютное время;
r — единица биологического (системно характерного)времени;
Ssp — специфическая энтропия одного вида живого,
е — основание натурального логарифма.
Важно здесь то, что минимум энтропии возникает при неравномерном распределении веществ в системе. Человеческая деятельность нарушает эту неравномерность, делает живое вещество гомогенным или даже, образно говоря, сдирает «живую кожу» с лика Земли, видоизменяет энтропийные и негэнтропийные процессы.
Антропогенное воздействие на окружающую среду оказалось деструктивным. Эволюция вынуждена идти экстенсивно, под воздействием внешних факторов, с темпом, диктуемым не ходом естественных явлений, а трансформацией природы человеком. Закон исторического развития биосистем работает не в полной мере или совсем не работает в силу того, что роль биотического воздействия на среду относительно снизилась. Преобладает преобразующая деятельность человека (рис. 12.45).
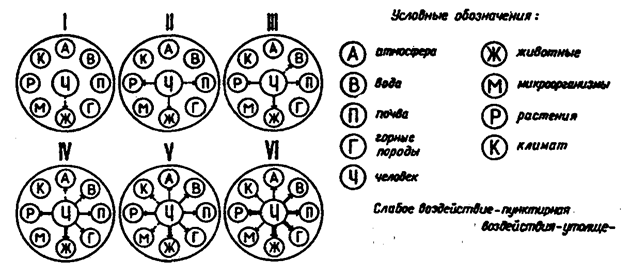
Рис. 12.45. Воздействие человека на природу на разных
стадиях развития производства:
I — период до времени использования огня; II — период со времени использования огня, появления и совершенствования примитивных орудий труда (100—10 тыс. лет до н. э.); III — период становления и развития земледелия и скотоводства (10 тыс. лет до н. э. — XIV в.); IV — период развития ремесел, появления и роста мануфактуры, расширения сельскохозяйственного производства (XV—XVIII вв.); V — период машинной индустрии, развития различных отраслей хозяйств (XIX в. — 1-я половина XX в.); VI — период научно-технической революции (2-я половина XX в.)
Здесь вслед за прямым уничтожением видов следует ожидать самодеструкции живого. Этот процесс фактически и идет в виде массового размножения отдельных организмов, разрушающих сложившиеся экосистемы. Насколько такое положение опасно для биосферы? Все зависит от темпов изменений. Следует учесть, что эволюция биосферы не была равномерной (рис. 12.46), и, несмотря на увеличение степени совершенства биогеохимического круговорота, этот процесс не шел гладко.
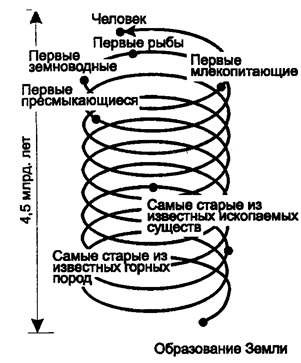
Рис. 12.46. Спираль времени
Закрученная в спираль лента графически изображает 4,5 млрд лет истории Земли. Отсчет времени начинается (нижний конец ленты) с формирования нашей планеты. Стрелками указаны ключевые моменты в эволюции жизни, обнаруживающие относительную молодость млекопитающих как целостной группы, не говоря уже о человеке, время существования которого - 1 млн лет или около того – указано на самом верху ленты. Одно деление нанесенной на ленте шкалы соответствует приблизительно 4,5 млн лет
Сегодня известны эволюционные катастрофы, происшедшие на нашей планете. Например, 650 млн лет назад эволюционно-экологический кризис привел к «внезапному» исчезновению многих видов одноклеточных водорослей. На рубеже 450 млн лет назад вымерло большинство панцирных обитателей океана, 230 млн лет назад исчезли многие виды гигантских амфибий, и, по эволюционным меркам, сравнительно быстро — 65 млн лет назад—вымерли гигантские рептилии и многие виды других групп организмов. Взгляды ученых на вымирание живых организмов неоднозначны. Так, вымирание гигантских рептилий связывают с похолоданием, прошедшим на Земле в результате падения огромного метеорита-астероида (предполагается, что он образовал крупнейший метеоритный кратер на территории современной Мексики). Похолодание привело к срыву инкубации яиц рептилий, а также могло дать преимущества для эволюции ночных групп животных и привести к исчезновению особо теплолюбивых дневных млекопитающих. Конечно, причины вымирания, его механизмы могли быть и другими. К едва заметным на взгляд человека, энергетическим перестройкам ведут даже незначительные изменения абиотической среды. Одно несомненно — вид никогда не исчезает один, всегда наблюдается изменение пищевых и информационных сетей. Происходит глобальная перестройка на всех уровнях экосистем. Одни виды исчезают, другие их замещают. Это явление находит отражение в правиле (принципе) катастрофического толчка: глобальная природная или природно-антропогенная катастрофа (сближение Земли с другим крупным космическим телом, столкновение с астероидом, резкое изменение климата, обеднение биоты и т. д.) всегда приводит к существенным эволюционным перестройкам, которые относительно прогрессивны для природы (адаптируют ее системы к новым условиям среды), но не обязательно полезны для вида или иной систематической категории, в том числе для человека и его хозяйственной деятельности. В связи с тем что отмечаются ускорения и замедления эволюции, действует и принцип прерывности и непрерывности развития биосферы: процесс медленного эволюционного изменения организмов закономерно прерывается фазами бурного развития и вымирания практически без переходных (палеонтологических) форм. Здесь интересен не столько механизм эволюции, а сам факт различного ускорения эволюционных процессов и их направленности. В случае, если ведущую роль в отборе играли верхние уровни природных систем и они же фактически направляли эволюцию, то антропогенные изменения биосферы, которые идут с большей скоростью, могут для нового ускорения эволюционных перестроек дать толчок в любой момент, вследствие чего произойдет капитальная перестройка экологических условий на Земле. Человечество как биологическое и социально-экономическое образование к таким преобразованиям едва ли готово. Нужны хотя бы общие показатели-рамки для установления, что является опасным, а что еще не грозит опасностью в ходе эволюции среды и жизни. По мнению ряда ученых, такими критическими показателями могут быть точки Пастера и правила одного и десяти процентов. Как известно из раздела 2.3, основной точкой Л. Пастера является момент, когда уровень содержания кислорода в атмосфере Земли в процессе эволюции составил 1% современного. Аэробная жизнь стала возможной с этого времени, что соответствует геохронологически архею. Считается, что накопление кислорода шло взрывообразно, в течение около 20 тыс. лет (рис. 12.47).
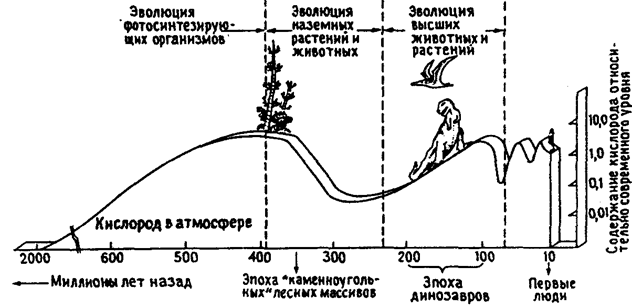
Рис. 12.47. Происхождение кислорода в атмосфере (по Е. Одуму, 1971)
Вторая точка Пастера — достижение также в архее содержания кислорода в атмосфере Земли около 10% современного. Создались предпосылки формирования озоносферы (рис. 12.48). Появилась возможность жизни на мелководьях, а в дальнейшем и на суше.
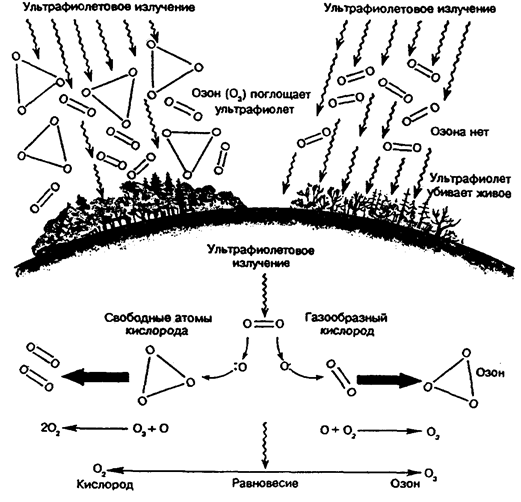
Рис. 12.48. Механизмы образования озонового слоя (внизу)
и его роль в атмосфере (вверху), по Е. А. Криксунову и др., 1995
Точки Пастера, как и закон пирамиды энергий Р. Линдемана (раздел 12.7), дали основание для формулировки правил одного и десяти процентов, получившего название закона Линдемана. Так называемое «магическое число» 1% возникает из соотношения возможностей потребления-энергии и «мощностей», необходимых для стабилизации среды. Доля возможного для биосферы потребления общей первичной продукции не превышает одного процента, что следует и из закона Р. Линдемана: около 1% чистой первичной продукции в энергетическом выражении потребляют позвоночные животные как консументы высших порядков, около 10% — беспозвоночные животные как консументы низших порядков, оставшуюся часть — бактерии и грибы-сапрофаги. Человечество, как только на грани XIX—XX вв.стало использовать большее количество продукции биосферы (в 90-х гг. XX в. — не менее 10%), так и перестал удовлетворяться принцип Ле Шателье-Брауна (примерно с 0,5% общей энергетики биосферы), рис. 12.49 и 12.50.
Рис. 12.49. Биосфера и человек-модель развития системы
их взаимоотношений (по Н. Ф. Реймерсу, 1990)
Примечание: толщина стрелок внутри кругов отражает силу воздействий; 1—3 — фазы преобразования природы человеком (промышленная, скотоводческо-земледельческая, индустриальная)

Принцип Ле Шателье-Брауна гласит, что при внешнем воздействии, выводящем систему из состояния устойчивого равновесия, равновесие смещается в том направлении, при котором эффект внешнего воздействия ослабляется. Так, растительность не давала прироста биомассы в соответствии с увеличением концентрации СО2 и т. д. Прирост связанного растениями углерода наблюдался лишь в XIX в.
Достаточно признанным считается порог потребления 5—10% суммы вещества, который приводит с переходом через него к заметным изменениям в системах природы. Он в большей степени принят на эмпирическо-интуитивном уровне, без различия форм и характера управления в данных системах. Приближенно намечающиеся переходы для природных систем можно разделить на с организменным и консорционным типом управления и популяционных систем. Для первых порог выхода из стационарного состояния до 1 % от потока энергии или «нормы» потребления и порог саморазрушения составляет около 10% этой «нормы». Для популяционных систем превышение в среднем 10% объема изъятия приводит к выходу этих систем из стационарного состояния.
Обратим внимание на формулировку «выход из стационарного состояния». Для глобальной энергетической системы такой выход, вероятно, происходит в рамках 0,1—0,2% возмущения общепланетных процессов, т. е. значительно раньше, чем наступают сбои в действии принципа Ле Шателье-Брауна и заметные природные аномалии. В подтверждение вышесказанного можно указать на то, что опустынивание начало существенно расти еще в прошлом веке. Трудно доказать или опровергнуть антропогенность климатических процессов, происходящих в последние два столетия.
Эволюционные переходы в биосфере занимают относительно небольшое время. Правила усиления интеграции биологических систем И. И. Шмалъгаузе-на гласят, что в процессе эволюции биологические системы становятся все более интегрированными, со все более развитыми регуляторными механизмами, обеспечивающими такую интеграцию. Н. Ф. Реймерс в работе «Системные основы природопользования» указывал на то, что разрушение более трех уровней иерархии экосистем абсолютно необратимо и катастрофично. Для поддержания надежности биосферы обязательна множественность конкурентно взаимодействующих экосистем. Таким путем шла эволюция биосферы. Антропогенные же воздействия нарушают этот ход. Правило множественности экосистем вытекает и из правила экологического дублирования, и вообще из теории надежности. Здесь интеграция оказывается «скользящей» по иерархической лестнице экосистем.
Дата добавления: 2015-12-01; просмотров: 1218;