Регуляция синтеза белков с точки зрения молекулярной биологии.
Процесс биосинтеза любой белковой молекулы в клетке включает в себя два основных этапа – транскрипцию (считывание информации с ДНК путем синтеза м-РНК) и трансляцию (синтез белка на м-РНК). Ключевую роль в обоих процессах играют играют молекулы нуклеиновых кислот, которые являются переносчиками информации в клеточных процессах. Аминокислотная последовательность первичной структуры любой белковой молекулы закодирована в нуклеотидной последовательности того или иного участка ДНК называемого геном. Следовательно регуляция синтеза белковых молекул это не что иное как регуляция транскрипции и трансляции (экспрессии) соответствующих им структурных генов. Для примера рассмотрим регуляцию экспрессии генов у прокариот.
Механизм генетической регуляции процесса индукции ферментов был расшифрован в экспериментах на кишечной палочке при изучении синтеза упоминавшегося фермента утилизации лактозы- β-галактозидазы. В 1961 г. F. Jacob и J. Monod на основании результатов генетического и биохимического изучения процесса утилизации лактозы бактериями E.coli К сформулировали концепцию, получившую широкую известность как «модель оперона».
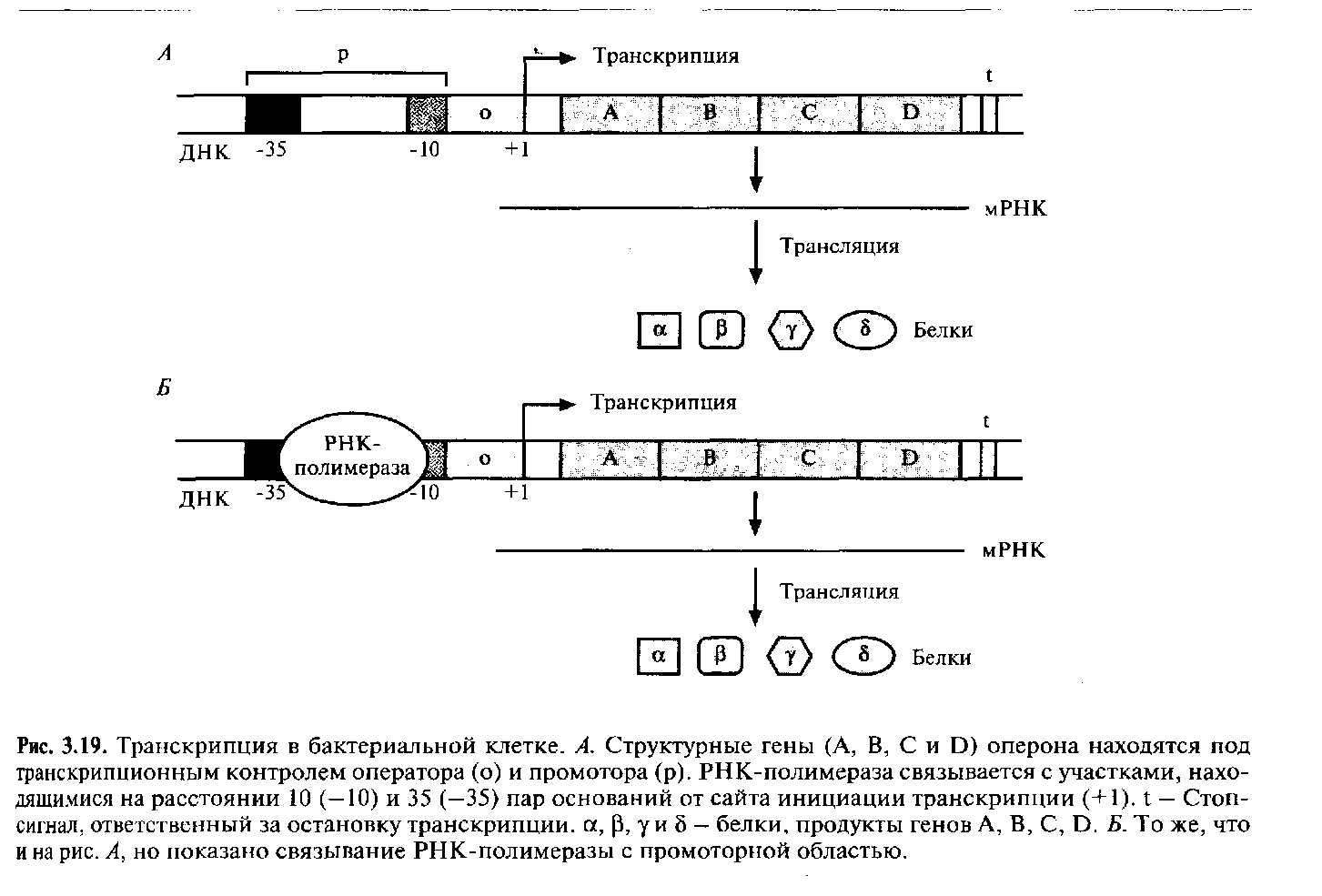
Согласно этой концепции, часто у бактерий белки-ферменты одного метаболического пути кодируются смежными структурными генами. Нуклеотидная последовательность, в которой закодировано более одного белка, называется опероном. Обычно оперон находится под контролем единственного промотора, и при его транскрипции образуется одна длинная молекула мРНК, кодирующая несколько белков. Промотор- это участок молекулы ДНК, с которым связывается РНК-полимераза, что сопровождается инициацией транскрипции соответствующих генов. При трансляции такой мРНК, в которой стоп-кодон последовательности, кодирующей один белок, соседствует со старт-кодоном гена следующего белка, синтезируется набор дискретных белков.
В большинстве структурных генов Е. coliимеются два сайта связывания для РНК-полимеразы. Один из них обычно представляет собой нуклеотидную последовательность
ТАТААТ (ТАТА-бокс, или бокс Прибноу),
АТАТТА
а другой - TTGAC
AACTG.
ТАТА-бокс и последовательность TTGAC расположены за 10 (область -10) и 35 (область -35) нуклеотидов до сайта инициации транскрипции соответственно (нуклеотид +1) (рис. 3.19). Обычно от участка между ТАТА-боксом и нуклеотидом +1 во многом зависит, будет ли происходить транскрипция данного оперона. В зависимости от способа регуляции транскрипции оперона этот участок называется оператором или активатором.
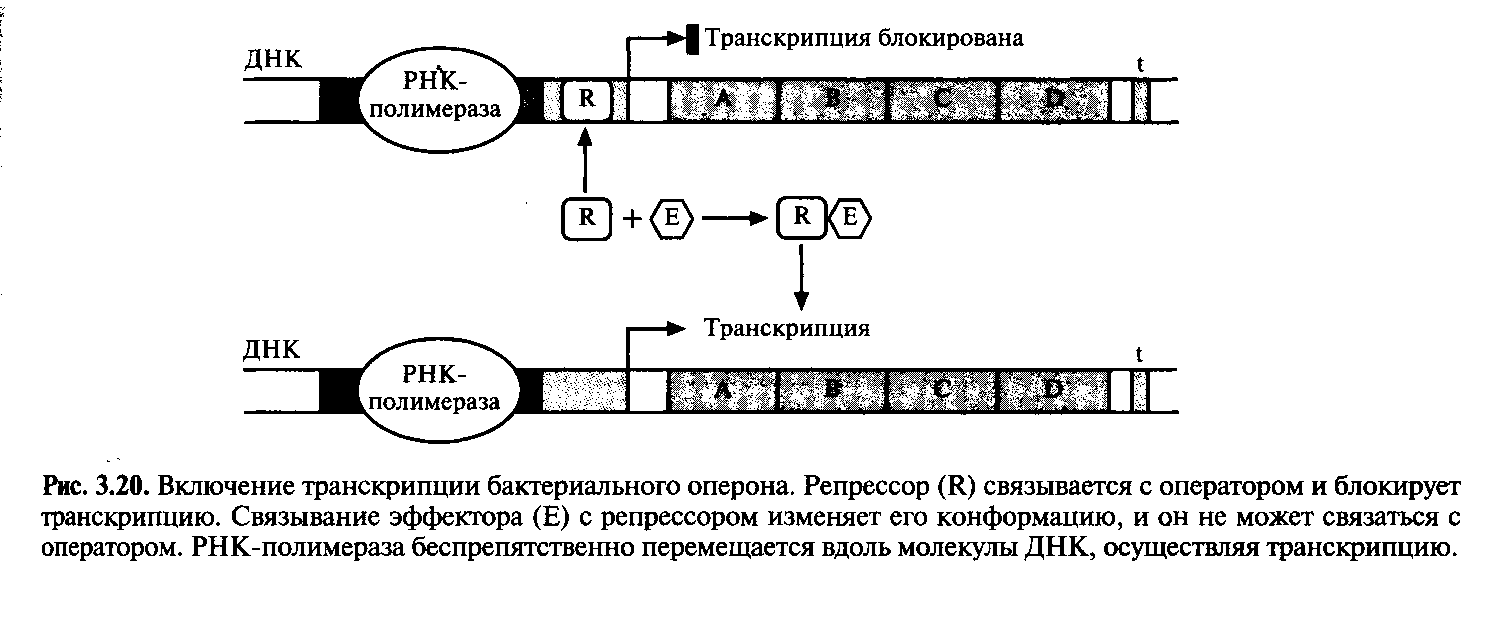
Для включения и выключения разных оперонов в ходе эволюции сформировалось множество регуляторных систем. Например, с операторной областью может быть связан регуляторный белок, называемый репрессором; он мешает перемещению РНК-полимеразы вдоль молекулы ДНК, и транскрипция блокируется (рис.3.20). Такой репрессор является аллостерическим белком, обладающим двумя специфическими центрами, один из которых характеризуется сродством к нуклеотидной последовательности области оператора, а другой – к молекуле эффектора.
Однако если с репрессором свяжется некое низкомолекулярное вещество (эффектор), то его конформация изменится таким образом, что его связывание с операторной областью станет невозможным, и транскрипция возобновится. Обычно эффектор разрушается клеточными ферментами. Когда его концентрация снижается, репрессор связывается с операторным участком, и транскрипция вновь прекращается. Операторный участок специфичен для каждого оперона, а эффектор взаимодействует только с определенным репрессором.
В качестве иллюстрации рассмотрим такой пример. Предположим, что клетка способна метаболизировать определенный сахар. Тогда синтез ферментов, расщепляющих этот сахар, будет бесполезной тратой клеточных ресурсов, если он отсутствует в среде. С другой стороны, если этот сахар имеется в достаточном количестве и является единственным источником углерода, то ферменты, отвечающие за его утилизацию клеткой, становятся совершенно необходимыми. В этом случае сахар действует как эффектор, препятствуя связыванию репрессора с операторным участком и таким образом обеспечивая транскрипцию оперона и синтез ферментов. При истощении запасов сахара в среде репрессор связывается с операторным участком и транскрипция оперона прекращается.
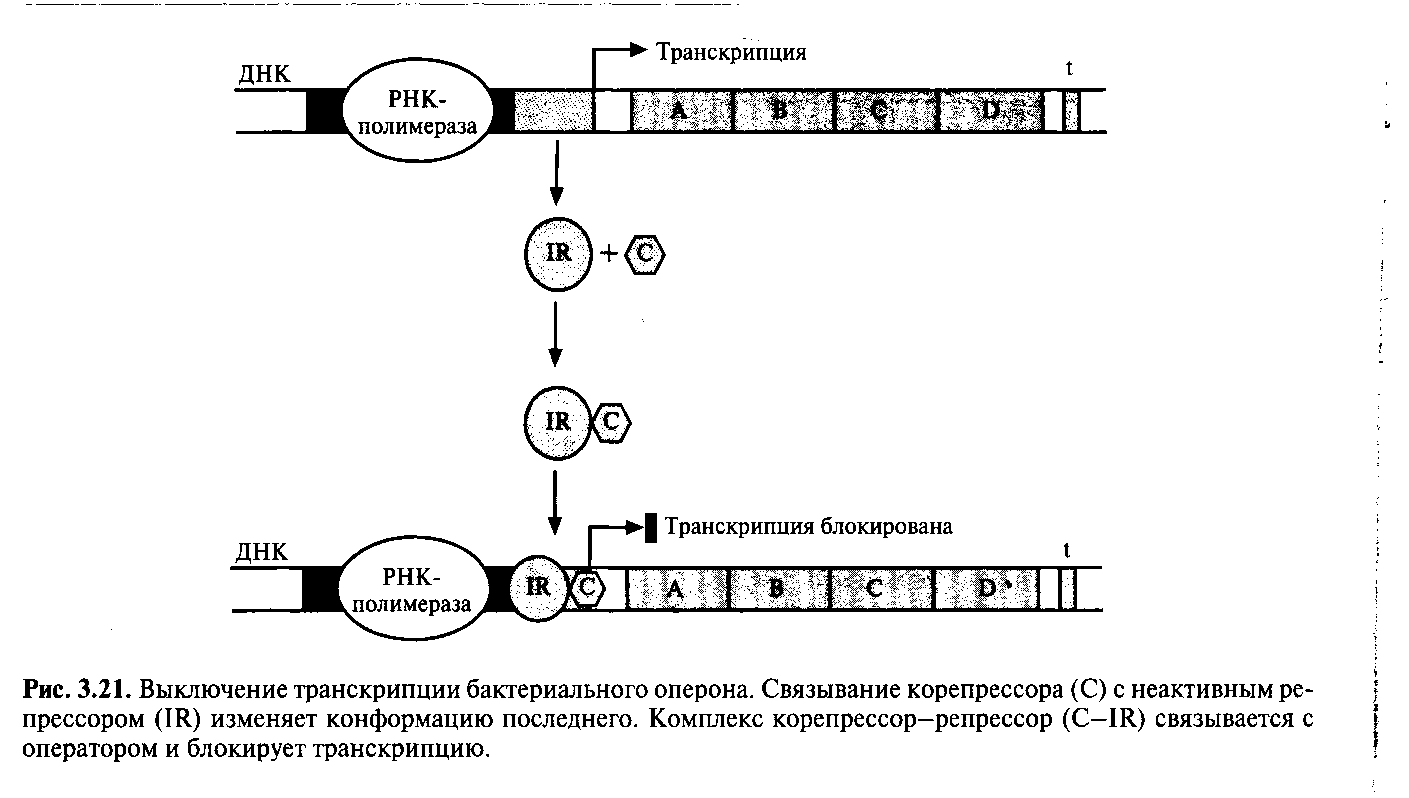
Нормальным состоянием других оперонов может быть состояние, при котором осуществляется их транскрипция, поскольку репессорный белок неактивен. В этом случае специфический эффектор (корепрессор), связываясь с неативным репрессором, вызывает в нем такие конформационные изменения, которые обеспечивают связывание комплекса с операторным участком, и транскрипция оперона выключается (рис.3.21). Сам по себе репрессор не способен связываться с оператором, поэтому при уменьшении концентрации корепрессора транскрипция возобновляется.
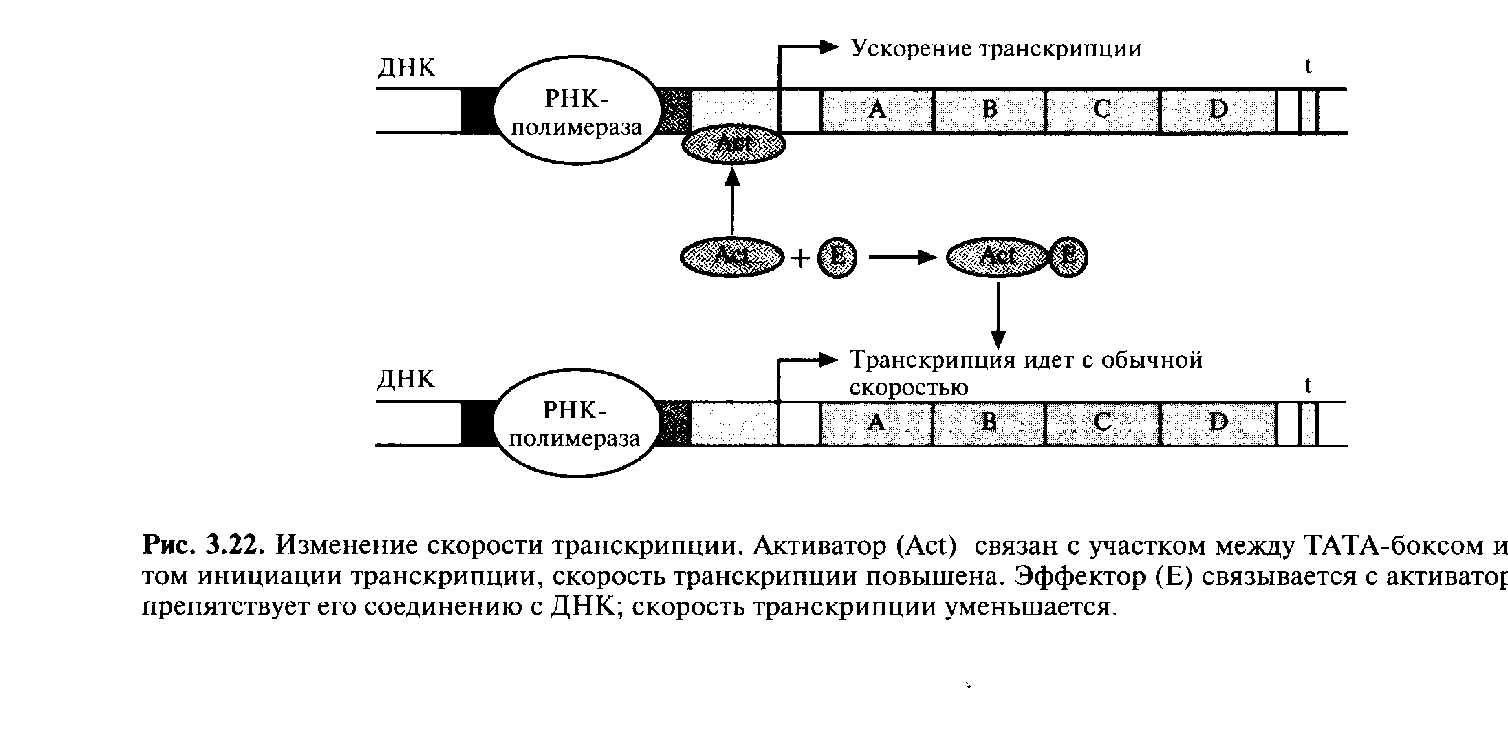
Регуляция транскрипции с помощью репрессора называется отрицательной. Если же система регуляции направлена на повышение скорости транскрипции, то она называется положительной. Рассмотрим вкратце этот процесс. Белок – активатор связывается с участком между ТАТА-боксом и сайтом инициации транскрипции. При это он не только не блокирует перемещение РНК-полимеразы вдоль молекулы ДНК, а напротив, ускоряет его, действуя как своего рода «смазка». Активаторы специфичны для определенных сайтов активации. Иногда с работающим активатором связывается эффектор, переводящий его в неактивную форму; тогда скорость транскрипции уменьшается (рис. 3.22). В других случаях эффектор, напротив, активирует неработающий активатор.
Следует отметить, что структурные гены одного метаболического пути не всегда объединены в единый оперон (наподобие лактозному), однако это не мешает их регуляции с помощью индукции или репрессии. Так, например, гены E.coli, детерминирующие структуру ферментов, обеспечивающих биосинтез аргинина, располагаются в различных областях хромосомы, но все контролируются одним и тем же геном-регулятором. Такая система образует регулон. Другим показательным примером является SOS-регулон, гены которого детерминируют структуру более десятка различных белков и ферментов, участвующих в репарации повреждений ДНК клетки. Все эти структурные гены регулируются одним репрессором – продуктом гена lехА. Опероны и регулоны, контролирующие взаимосвязанные физиологические функции обнаружены у всех генетически изученных видов бактерий.
Лекция № 3
Дата добавления: 2016-01-30; просмотров: 2784;