Методы феногеографии
Главный метод феногеографии – сопоставление фенофондов. Его можно провести двумя путями – на карте и без нее. В обоих случаях анализ производится двумя способами: по одному фену либо сразу по комплексу фенов. Часто сопоставление по одному фену предшествует последующему анализу комплекса фенов, но порой оно может представлять самостоятельный микроэволюционный интерес.
В случае сопоставления по одному фену на карту наносится либо концентрация данного фена в разных точках исследования, либо просто отмечается факт присутствия данного фена в этой части ареала. В зависимости от задач исследования и то и другое изображение может оказаться одинаково удобным.
На рис. 18 показано изученное Н. В. Тимофеевым‑Ресовским распространение фена, определяющего слияние пятен на надкрыльях растительноядной божьей коровки. Видно, что этот фен занимает большую часть внутри ареала вида и обладает как бы собственным ареалом. При таком подходе нас интересует сам факт распространенности фена в пределах ареала вида, а не та или иная его частота. На таких картах можно обнаружить адаптивные границы распространенности фенов, найти корреляции в распространении фена с какими‑либо факторами среды и т. д.
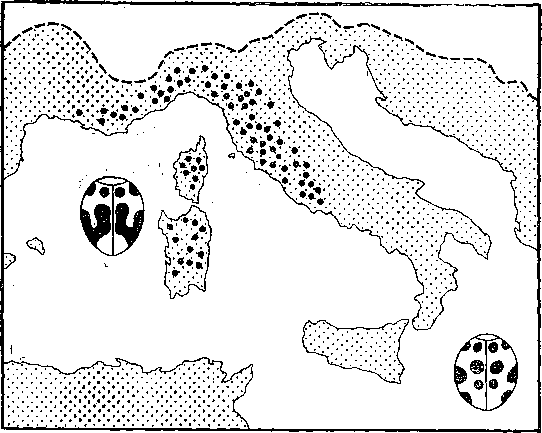
Рис. 18. Распространение фена слияния пятен на надкрыльях (крупные черные точки) у растительноядной божьей коровки Epilachna chrisomelina в Средиземноморье
Примером подобной работы может служить исследование по распределению фена меланизма у хомяков на Украине, выполненное в конце 30‑х годов известным советским генетиком С. М. Гершензоном. Ученому удалось выяснить, что распространение меланизма связано с повышенной влажностью местообитаний. Однако чаще, чем факт простого присутствия или отсутствия данного фена в том или ином участке ареала, в исследованиях анализируется распространенность этого фена. Выше упоминался широко известный в мировой литературе случай распространения мутации (фена) simplex в строении зубов обыкновенной полевки в Средней Европе. Из зоны с повышенной концентрацией этого фена в Ютландии во все стороны происходит как бы растекание признаков с постепенно понижающейся концентрацией этого фена. На рис. 19 показано распределение в пределах ареала прыткой ящерицы фена прерывистой полосы по центру спины. Видно, что зоны повышенной концентрации фена довольно четко отделены от зон пониженной концентрации. Проявляются как будто определенные границы его пониженной или повышенной концентрации. На аналогичных картах, составляемых для прыткой ящерицы по другим фенам, есть очень интересные границы: для одних фенов р. Волга, например, является непреодолимой преградой; другие фены в высокой концентрации встречаются на ее западном берегу и в очень низкой концентрации – на восточном. Для некоторых же фенов как будто и не существует такой преграды, как Волга.
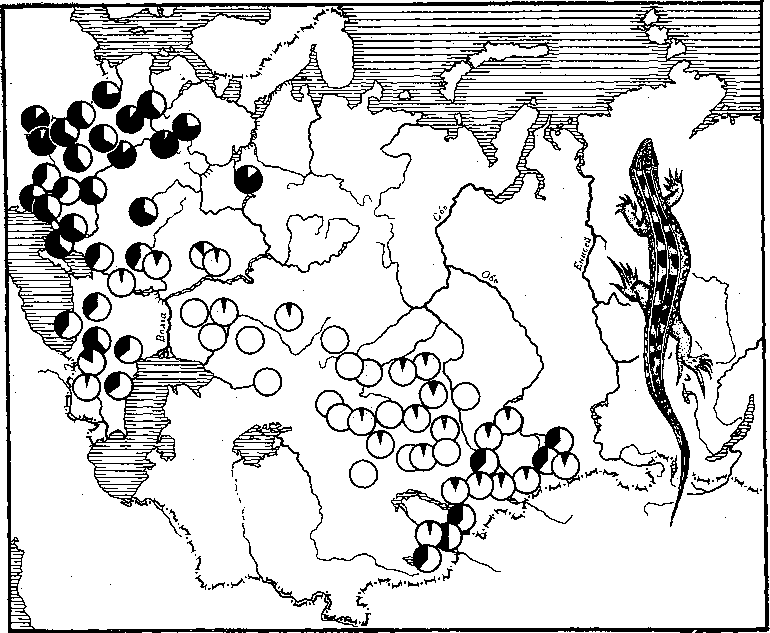
Рис. 19. Распределение фена «прерывистая полоса на спине» (черный сектор) на большей части ареала прыткой ящерицы
Выше не раз подчеркивалось, что не всякий анализ единственного фена окажется содержательным и интересным уже потому, что каждый фен обладает определенным масштабом, в котором и проявится его закономерное распределение. Наглядным примером служит распределение фена желтой окраски раковины у виноградной улитки, вскрытое в работах английского генетика Дж. Джонса в 1974–1978 гг. Оказалось, что в масштабе участка протяженностью в пять километров распределение желтых раковин случайно, в масштабе территории площадью 22 500 км2 также не прослеживается каких‑либо закономерностей. Случайным оказывается и распределение фена в масштабах всей Великобритании. Однако в масштабе Европы распределение фена желтой окраски оказывается клинальным, т. е. подверженным отбору и явно адаптивным.
Прежде чем рассмотреть пути сопоставления фенофондов по многим фенам, надо сделать одно отступление в область инженерной психологии. Выше говорилось о том, что оптимальным для одновременного восприятия человеческим глазом является число разнородных знаков, не превышающее девяти. Это обстоятельство надо иметь в виду при нанесении на карту нескольких фенов одновременно. Лучше всего, если на карте будет не более семи разных значков, иначе карта будет трудно читаема.
Оказывается, что разные методы кодирования признаков также резко различаются по эффективности зрительного восприятия. Отдельные фены на карте можно обозначить цифрами, буквами или значками разной конфигурации (треугольник, квадрат, ромб, круг и т. д.), но можно, наконец, изображать фены различным цветом. Эксперименты показали, что для определения места какого‑либо фена на карте эффективнее всего использовать цвет.
При анализе одного или нескольких фенов их можно наносить на карту без учета степени концентрации, но наиболее распространено изображение нескольких фенов с учетом их концентрации. Концентрацию можно показать величиной значка, густотой расположения символов на карте или – если наносятся частоты признаков, в сумме составляющие 100 %,– вариантами секториальных графиков. Если же требуется дать сопоставление нескольких независимых признаков из разных групп фенов, несомненно, лучшим является метод изображения «розой ветров».
Последним большим направлением в методике фенографических исследований можно рассматривать вычисление обобщенного фенетического показателя. Наиболее распространено в современных зарубежных работах, пожалуй, сравнение отдельных популяций и групп по так называемым коэффициентам сходства. Полученные данные могут сравниваться непосредственно на карте, в таблицах или служить основанием для построения дендрограмм, изображающих фенетическое сходство исследуемых группировок. Этот метод дает очень хорошие результаты и заслуживает широкого распространения в отечественных работах.
Другой, пока мало распространенный метод сравнения по обобщенному показателю – простой метод балльного сравнения. Он успешно применяется в случаях выявления каких‑то промежуточных форм между уже известными двумя или более крайними формами. Характерный результат этого метода– выявление гибридных зон между подвидами, «полувидами» и видами. Сущность метода можно проиллюстрировать на примере обстоятельного исследования двух «полувидов» американской иволги, проведенного американским зоологом Дж. Райзингом.
На Атлантическом побережье Северной Америки обитает балтиморская иволга, на Тихоокеанском – иволга Буллока, по центральной части континента с юга на север проходит широкая гибридная зона между этими «полувидами». Признаки окраски были разбиты на девять отдельных групп (окраска лба, шеи, ушной области, горла и т. д.). В каждую группу входило от трех до пяти дискретных вариантов окраски – фенов, расположенных в порядке увеличения значения условного балла для каждого фена от признаков типичной балтиморской иволги, принимаемых за 0, до признаков, характерных для иволги Буллока. Выявляющаяся изменчивость суммарного балла окраски всех признаков показывает широкую зону гибридизации, носящую характер клинальной изменчивости и коррелирующую с величиной осадков. Аналогичные сопоставления можно делать и без карты, в виде таблицы.
* * *
Не случайно рассказ о феногеографии оказался самым длинным в этой книжке. Феногеография действительно является одним из самых главных разделов фенетики, так как позволяет ставить и решать наиболее сложные проблемы популяционного исследования, начиная от поиска популяционных границ и кончая восстановлением хода эволюции – изучением микрофилогенеза вида. Важное значение должна иметь феногеография для правильной организации многих промыслов и ряда биотехнических мероприятий. Знание точного распространения хозяйственно важных признаков в пределах ареала вида – надежная основа для планирования биотехнических мероприятий. Хороший пример такого рода – восстановление численности соболя в нашей стране: оно проводилось посредством отлова и выпуска в подходящие местообитания соболей из лучших кряжей (популяций и групп популяций, характеризующихся исключительно удачным набором хозяйственно важных признаков – темной окраской, густотой меха, крупными размерами зверьков).
Важное значение имеет феногеография и при решении главного вопроса внутривидовой систематики: целесообразно или нецелесообразно выделять какие‑то группы популяций в качестве отдельных подвидов.
Наконец, велико значение феногеографии для развития теории микроэволюции: изучения механизмов формообразования, пространственных тенденций в развитии внутривидовой изменчивости, образования центров многообразия, клинальной изменчивости и т. п.
Заключение
Причиной написания этой книжки послужило быстрое развитие нового направления исследований в области изучения природных популяций – фенетики. В самой общей форме фенетика – это применение генетических идей и методов при любом негенетическом изучении природных популяций (с позиций зоолога и ботаника, эколога и морфолога, физиолога и этолога). С одной стороны, фенетика позволяет ставить и решать популяционные задачи, которые раньше если и ставились, то не могли быть решены на достаточно обширном материале. С другой стороны, она позволяет ставить такие задачи и проблемы, которые раньше вообще не могли быть поставлены из‑за отсутствия методов их решения.
Фенетика позволяет разрешить противоречие между практической невозможностью широкого генетического исследования видов и методологической необходимостью получения таких данных. Фенетический подход позволит сравнительно быстро описать и расшифровать в первом приближении сотни и тысячи разнообразных эволюционных ситуаций в природе. Это выяснение реальных путей микроэволюции, видимо, и станет фундаментом теории управления эволюционным процессом.
Нет сомнения, что «надев генетические очки», но не ограничивая себя кругом проблем классической генетики, исследователи большинства разделов полевой биологии, а возможно, и ряда разделов экспериментальной биологии посредством использования фенетических приемов и методов получат новые возможности для решения научных проблем. Все это должно оказаться полезным в продвижении по пути решения кардинальной проблемы современной биологии – управления эволюционным процессом, без которого, как мы уже говорили в начале книги, невозможно представить будущее полноценное существование человечества в биосфере Земли.
Насколько продуктивным окажется фенетический подход в конкретном исследовании, зависит не только от используемых фенетических методов. Фенетика дает ключ к анализу происходящих в природе микроэволюционных событий, скрытых от кратковременных наблюдений с помощью обычных классических методов. Какая дверца будет открыта этим ключом – зависит от самого исследователя.
Широкая междисциплинарная основа фенетики, с одной стороны, и желательность – в связи с катастрофически растущим влиянием цивилизации на живую природу – скорейшего перехода от бесконтрольного вмешательства в ход эволюции живой природы на Земле к управляемой эволюции – с другой, заставляет писать о фенетике не только в трудах научных конференций и узкоспециальных монографиях, но и в издании, обращенном к широкому кругу читателей, заинтересованных в изучении живой природы.
Если эта книга привлечет внимание читателя к тем или иным проблемам эволюционной и популяционной биологии, задача автора будет выполнена наполовину. Если же она пробудит у читателя интерес к изучению дискретных альтернативных признаков в природных популяциях и вызовет желание у негенетиков «надеть генетические очки», а у генетиков – вспомнить подходы и методы, которые столь успешно начал развивать в своих трудах Н. И. Вавилов, и резко расширить круг изучаемых объектов – задача автора будет выполнена полностью.
[1]См. в кн.: Грегор Мендель, Шарль Нодэн, Огюстен Сажре. Избранные работы, М.: Медицина, 1968, с. 64.
[2]Там же.
[3]Цит. по: Гайсинович А. Е. Зарождение генетики, М.: Наука, 1967, с. 100.
[4]Цит. по: Гайсинович А. Е. Зарождение генетики, с. 127.
[5]Shull H. G. Genetic definitions in the New Standard Dictionary. – Amer. Natur., 1915, vol. 49, N 577, p. 59.
[6]Цит. по: Гайсинович A. E. Зарождение генетики, с. 142.
[7]Вавилов Н. И. Научные основы селекции пшениц. – Избр. произв. Л.: Наука, 1967, т. 2, с. 86–87.
[8]Там же, с. 228.
[9]Там же, с. 75.
[10]Там же, с. 41.
[11]Там же.
[12]Вавилов Н. И. Закон гомологических рядов в наследственной изменчивости. – Избр. произв. Л.; Наука, 1967, т. 1, с. 37–38.
[13]Серебровский А. С. Проблемы и методы геногеографии. – Тр. Всесоюз. съезда по генетике, 1929, т. II, с. 72.
[14]Борисяк А. А. Основные проблемы эволюционной палеонтологии. М.; Л.: Изд‑во АН СССР, 1947, с. 73.
[15]Тимофеев‑Ресовский Н. В., Яблоков А. В. Фены, фенетика и эволюционная биология, – Природа, 1973, № 5, с. 40–51; Тимофеев‑Ресовский Н. В., Яблоков А. В., Глотов Н. В. Очерк учения о популяции, М.: Наука, 1973, с. 278.
[16]Аллели – формы состояния гена, вызывающие фенотипические различия и располагающиеся в одинаковых участках гомологических (парных) хромосом.
[17]Кондаков Н. И. Логический словарь‑справочник. М.: Наука, 1975, с. 477.
[18]Отступая от хода изложения, следует сказать, что самая обычная методическая ошибка в любом творческом процессе состоит в том, что исследователи размышляют над «уравнением со многими неизвестными». Такие уравнения не имеют однозначных решений, поэтому попытки их точного решения бессмысленны. В любом исследовании надо стараться составить уравнение с одним или двумя неизвестными (уравнение с двумя неизвестными имеет ограниченное число решений, и над этим ограниченным числом решений иногда интересно подумать). Практический вывод из этого рассуждения прост: если в наших биологических конструкциях и гипотезах получается более чем два неизвестных, их необходимо упрощать и делать более строгими, доводя число неизвестных до одного‑двух. Иначе вся работа будет напрасна и определенный ответ получить будет невозможно.
[19]Это ясное в общих чертах положение может осложниться одним обстоятельством: в направленности можно уловить действие не современного отбора, а проявление результатов отбора, действовавшего много‑много поколений назад и сформировавшего определенные черты строения организмов. Эти черты неизбежно определяют некоторые возможные направления дальнейшего процесса отбора. Интенсивная разработка проблемы направленности эволюции показывает, что такое «последействие» отбора, зафиксированного в строении каждого живого существа, – одно из широко распространенных эволюционных явлений.
[20]От слова «панмиксия» – свободное скрещивание особей в пределах популяции (или другой группы).
[21]Клина – градиент значения какого‑либо признака внутри ареала вида.
Дата добавления: 2016-01-26; просмотров: 1100;