Несколько подробнее о фенах млекопитающих
Все сказанное выше относилось к общим принципам выделения фенов. Теперь же посмотрим с фенетической точки зрения только на одну‑единственную группу животных – на млекопитающих. В недалеком будущем, несомненно, атласы фенов по разным группам животных и растений окажутся необходимой принадлежностью многих лабораторий. Пока таких атласов нет, и приводимый ниже перечень вероятных (или уже обнаруженных) фенов в разных системах органов млекопитающих позволит составить более полное представление о возможностях фенетического подхода.
Ниже перечисляются группы признаков, наследственная обусловленность которых подтверждена либо экспериментально в генетических исследованиях, либо косвенно – данными о маркировании ими различных групп особей внутри вида.
Форма и размер тела. Главные дискретные вариации – карликовость и гигантизм, укороченные конечности (пример со знаменитой анконской овцой, приводимый в «Происхождении видов» Ч. Дарвином), «повернутый нос», резкое укорочение и удлинение морды, головы, короткохвостость и бесхвостость, потеря отдельных пальцев, слияние пальцев на передних и задних конечностях, добавочные пальцы. По форме тела сейчас можно выделить несколько десятков фенов.
Окраска. Шесть главных и много дополнительных генов влияют на цвет покровов млекопитающих, определяя появление многих десятков дискретных признаков по цвету. Не менее 10 генов связаны с пятнистостью и полосатостью. В целом на хорошо генетически изученных объектах (мышь, крыса, кролик) показано наличие нескольких сотен дискретных признаков окраски, которые могут считаться фенами (рис. 9). Среди них пятна разного цвета, оттенка, формы и размера: светлые на темном фоне, темные на светлом фоне, с резкими границами, расплывчатые, с цветной границей, мелкие и крупные, округлые и угловатые, сложной формы (крестообразные, сердцеобразные, лирообразные, звездочки, кольца, розетки и т. п.); окраска головы: черная, белая, светлый верх – темный низ («барсучий тип»), темный верх – светлый низ (есть порода овец, у которых сочетание двух последних типов окраски при скрещивании дает черноголовость), окраска типа маски («очки»); уши: темные, светлые, темные или светлые на кончиках; темная макушка, темные и белые пятна на щеках; белый клок волос на лбу; темные пятна около ушей, «звездочка» на лбу; поседение волос до наступления зрелости и т. д.
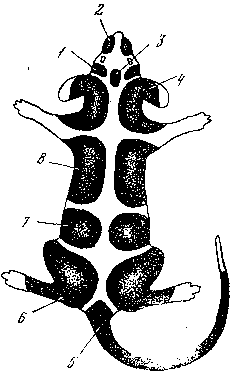
Рис. 9. Зоны действия отдельных генов и групп генов (1–8 ), определяющих появление разных вариантов окраски покровов млекопитающих
Строение волосяного покрова: локоны на спине, завитки шерсти на плечах и бедрах, отсутствие волос на определенных участках тела, очень длинные волосы, пучки волос на ушах, длинные волосы пучками по всему телу, «плюшевый» волосяной покров без остевых волос, волнистость, закрученные и изогнутые вибриссы – вот лишь некоторые из дискретных признаков, касающихся волосяного покрова черной крысы. Можно предположить, что по особенностям строения волосяного покрова у разных млекопитающих можно выделить около сотни фенов.
Производные кожного покрова. Вибриссы. По особенностям расположения и встречаемости вибрисс известны десятки вариантов у каждого из изученных в этом отношении видов. У крупных хищных расположение вибрисс – один из удобных признаков для индивидуальной идентификации.
Млечные железы. Известно много дискретных вариантов расположения и числа сосков млечных желез у млекопитающих, чешуйчатости хвоста у грызунов, папиллярных узоров у человека и приматов; варианты в расположении подошвенных бугорков и в папиллярных узорах носового зеркала у копытных.
В целом среди особенностей кожного покрова можно выделить наверное 500–700 фенов.
Скелет. Среди особенностей скелета можно найти немало фенов: множество мелких вариаций черепа (сотни вариантов на достаточно крупных черепах), зубной системы, которую условно рассматриваем в этом разделе (в частности, у человека – пока наиболее изученной, хотя и не наиболее сложной – выделяется около 400 вариаций, множество из которых, видимо, является фенами; особенности строения зубов грызунов позволяют выделить десятки фенов), скелет переднего и заднего поясов конечностей, осевого скелета. Во всех участках скелета уже выделены сотни наследственных изменений. В целом среди наследственных особенностей скелета можно выделить не меньше нескольких сот фенов.
Пищеварительная система. Форма, число и расположение разного типа вкусовых сосочков на языке дают десятки вариантов. Дискретна изменчивость формы долей печени, поджелудочной железы, желчного пузыря и желчных протоков (вплоть до отсутствия пузыря – признак, обычный для внутривидовой изменчивости у многих видов). Варьируют граница между роговым и железистым эпителием передней и задней кишки, форма складчатости железистого эпителия на разных участках кишечной трубки и т. п. Пищеварительная система сравнительно мало изучена с фенетической точки зрения, но можно предполагать возможность выделения 100–150 фенов.
Мускулатура. Внутривидовая изменчивость мышечной системы практически не изучена, однако все без исключения исследования на достаточно большом материале показали удивительное разнообразие в строении и топографии даже самых обычных и хорошо развитых мышц. В будущем возможно выделение многих десятков фенов.
Дыхательная система. Дольчатость легких (на примере популяции котиков показано, что эти фены пригодны для межпопуляционного уровня), ветвление бронхиального древа, легочного артериального древа, строение дивертикулов гортани и носовых проходов показывают наличие дискретных вариаций. В дальнейшем возможно выделение многих десятков фенов.
Мочеполовая система. Дискретные варианты топографии почек и надпочечников (в случае дольчатых почек для особенностей объединения отдельных долек), впадения мочеточников, рисунка эпителия, выстилающего мочевой пузырь, формы и расположения os penis и os clitoridis, складчатости влагалища, формы семенников и яичников. В этой системе можно выделить многие десятки фенов.
Нервная система и органы чувств. Известны дискретные варианты окраски и аномалии строения глаз (не менее 20–25 различных фенов), строения ушной раковины (десятки фенов только у человека); варианты топографии отдельных нервных стволов и сплетений (все без исключения исследованные серии показывают огромную изменчивость). Нервная система мало изучена фенетически, и пока возможно выделение нескольких десятков фенов.
Кровеносная система. Известно огромное число наследственных вариантов топографии крупных и мелких кровеносных стволов (артерий и вен) в разных частях тела; изменчива форма и топография селезенки. Кровеносная система наряду с системами органов кожного покрова и скелета – одна из наиболее перспективных с позиции фенетического исследования и потенциально содержит сотни сравнительно легко обнаруживаемых фенов.
Кариотип. Большое число вариантов известно для строения хромосомных наборов: внутривидовой хромосомный полиморфизм характерен, как показывают многочисленные исследования, видимо, для всех без исключения видов млекопитающих.
В общей сложности у млекопитающих сейчас известно не менее двух‑трех тысяч наследственно детерминированных морфологических дискретных признаков – фенов. Несомненно, с развитием фенетического подхода число этих признаков будет стремительно расти и может составить тысячи для каждой системы органов.
О реальной возможности такого резкого увеличения числа фенетических признаков говорит ситуация, сложившаяся с биохимическими признаками («биохимическим фенотипом», как все чаще совершенно точно называются картины, получаемые при биохимическом исследовании методами электрофореза, иммуннологическими и другими). Еще 30–40 лет назад были известны единичные биохимические признаки, подверженные внутривидовой изменчивости, сегодня известны многие сотни таких признаков у многих видов.
Большое число фенов должно быть обнаружено при популяционно‑физиологических исследованиях. Вот несколько из множества подобных примеров. В результате направленной селекции получены линии крыс с высоким и низким кровяным давлением; эти различия связаны с вариациями одного гена, контролирующего синтез кортикостерона. Известны дискретные генетические вариации в содержании гормонов роста, гонадотропинов, инсулина, вазопрессина, катехоламина, простагландина и многих других гормонов у целого ряда видов млекопитающих. Сейчас такие исследования ведутся все шире, и, по‑видимому, в области физиологии можно в ближайшие годы ожидать выделения нескольких десятков, а может быть, и сотен фенов.
Значительно больше фенов уже сегодня выделяется в поведении млекопитающих. Развитие этологии показало такое разнообразие мельчайших наследственных вариантов поведения, о котором еще сто лет назад даже трудно было предполагать. (Впрочем, всем хорошо известны врожденные мельчайшие особенности человека, передающиеся из поколения в поколение, – походка, характер жестов и т. п.). Все больше накапливается данных по фенам голоса млекопитающих.
Итак, если к упомянутым выше группам морфологических фенов добавить биохимические, физиологические и этологические фены, окажется, что сегодня в распоряжении исследователей имеются тысячи точных признаков. Выбор нескольких десятков из них для конкретного популяционного исследования сейчас возможен практически для любого из видов млекопитающих.
Адаптивен ли фен?
В той или иной форме этот вопрос, вынесенный в заголовок раздела, встает почти всегда при рассказе о работах в области фенетики. Выше уже шла речь о фене левозакрученности у одного из видов наземных моллюсков. Оказалось, что особи, несущие этот фен, отличаются по характеру потребления энергетических запасов. Это говорит о том, что левозакрученность – не нейтральный признак и находится под контролем естественного отбора. Недавно свердловские зоологи показали, что различия между бесполосыми и полосатыми формами у других видов того же рода четко соответствуют различиям по интенсивности выделения CO2 при дыхании.
Работ подобного рода можно привести много, и все они позволяют прийти к общему заключению: во всех специально исследованных случаях особи – обладатели разных фенов всегда обнаруживали те или иные адаптивные различия. Фен красной окраски у двуточечной божьей коровки каким‑то образом связан со способностью лучше переносить холод во время зимовки, а фен черной окраски – с более интенсивным размножением летом. Фен желтой окраски у калифорнийской полевки связан с лучшей выживаемостью при низкой температуре, а также с лучшим размножением при питании зеленым кормом (а не семенами), чем у особей с типичной серой окраской. Фен серой окраски у американской лягушки‑сверчка связан с устойчивостью организма к потере влаги и лучшей сопротивляемостью некоторым инфекциям и т. д.
Принцип плейотропии – действие одного гена на многие признаки – означает, что между особями, несущими разные фены, должны существовать многочисленные различия в биологических признаках и свойствах. Наверное, некоторые фены могут иметь и непосредственное адаптивное значение. В других случаях фены имеют косвенное адаптивное значение, будучи связанными в развитии организма с какими‑то важными биологическими признаками и свойствами.
Косвенным, но достаточно определенным подтверждением адаптивного характера фенов являются данные по сочетаниям различных дискретных вариантов структуры жевательной поверхности зубов рыжей полевки. Пять вариантов строения третьего верхнего и четыре варианта строения первого нижнего коренных зубов этого вида, выделенные саратовскими зоологами, теоретически могли бы дать 400 сочетаний. Однако в большой серии изученных особей было найдено немногим более 10 % от этого, числа. Если бы выделенные варианты строения были безразличны для организма, то можно было бы ожидать самых разных сочетаний, Но это не так, и, видимо, большая часть сочетаний оказывается неблагоприятной для организма.
Другой важный показатель адаптивной «небезразличности» фенов – удивительное постоянство их частоты в природных популяциях. Сравнение частоты некоторых фенов рисунка спины у прыткой ящерицы в сборах, произведенных в одном и том же месте, но с промежутком в 70 лет, показало ее стабильность. Есть много других примеров сохранения постоянной частоты фенов (линейных различий) на протяжении десятков поколений у многих видов дрозофил, лабораторных мышей, кроликов, крыс и практически любых лабораторных животных и других животных, разводимых в неволе, равно как среди культурных растений и домашних животных. Если бы фены, характеризующие все эти генетически более или менее однородные совокупности особей, не имели адаптивного значения, то трудно было бы представить себе их поддержание в стабильном состоянии даже на протяжении немногих поколений.
Важно подчеркнуть, что незнание конкретного адаптивного значения данного фена не может служить препятствием для его использования в качестве признака – маркера природных группировок особей. Этот вывод методически важен, так как позволяет не отвлекаться на изучение конкретных адаптаций, которые, кстати говоря, никогда не могут быть изучены полностью. Этот вывод, звучащий, может быть, несколько неожиданно, неизбежно следует из ограниченности нашего познания в любой данный момент времени. Ограничусь только одним примером.
Долгое время считалось, что крылья летучих мышей являются только органом полета. Но вот точные наблюдения в природе показали важную роль крыльев у летучих мышей как сачка во время ловли добычи. Вскоре выяснилась огромная роль крыльев летучей мыши в терморегуляции: это единственные большие не покрытые мехом участки тела и они значительно охлаждаются во время полета. С другой стороны, животное может таким образом складывать крылья во время отдыха, что образуется как бы дополнительный кожистый термоизоляционный покров. Итак, у крыльев летучей мыши есть функция полета, сачка, разнообразные функции терморегуляции. Казалось бы, и все. Но физиологи напомнили, что кожа млекопитающих – это орган, в котором происходит синтез витамина D, и этот процесс идет лишь под влиянием солнечного света. Может быть, большие крылья с их огромной поверхностью кожи в какой‑то степени компенсируют летучим мышам недостаточную выработку витамина D, ведь солнечный свет обычно не попадает на них, поскольку все эти животные сумеречные или ночные. В будущем наверняка выяснятся еще какие‑то функции их крыльев. И этот процесс углубления познания будет идти бесконечно, параллельно нарастанию суммы знаний о строении и функционирований всего живого вообще.
Ситуация, описанная в примере с летучими мышами, иллюстрирует принцип мультифункциональности любого органа или структуры – заведомого наличия множества функций у каждого органа или части тела в организме любого существа. Но раз это множество никогда не будет исчерпано нашим познанием, не ошибочным ли (точнее, нужным ли) является требование предварительно выяснить адаптивное значение того или иного фена и лишь после этого использовать такой признак в исследованиях?
Для изучения природных совокупностей особей с помощью дискретных признаков‑фенов нам достаточно общего знания того, что любой признак организма – и фен в том числе – должен иметь то или иное непосредственное или опосредованное адаптивное значение.
* * *
Теперь, когда читатель знаком и с примерами фенов, и с основными свойствами фенов как дискретных, наследственно обусловленных признаков, и с особенностями выделения фенов, нужно, наконец, рассказать о том, что же можно выяснить в природе с помощью фенов.
Дата добавления: 2016-01-26; просмотров: 1071;