Глава III. Что такое фен?
Выше уже говорилось, что фенетика – это распространение генетических подходов и принципов на виды и формы, генетическое изучение которых затруднено или невозможно. Предмет фенетики – внутривидовая изменчивость, доводимая в конечном итоге до рассмотрения дискретных альтернативных признаков особи – фенов. Методы фенетики заключаются в вычленении различных фенов, характерных для изменчивости изучаемых форм, количественное и качественное изучение фенов в популяциях и других внутри‑, а также и межвидовых группах особей. Цель фенетики – разработка вопросов микроэволюции, теоретической систематики, практической биотехнии и других проблем, связанных с популяционным исследованием видов в природе.
Для первого знакомства и для стимулирования дальнейшей разработки этого нового направления популяционной биологии рассмотрим последовательно лишь основные проблемы, возникающие при изучении фенов, фенофонда и феногеографии – трех основных разделов фенетики. Начнем с описания фенов.
Как выглядит фен
Фен – отдельный дискретный (резко отделенный от других), наследственно обусловленный признак индивида. Эта формулировка понятия фена не полная – более полная будет приведена несколько далее, после рассмотрения ряда примеров.
Фенами следует считать все те «мутантные признаки», «мутации», которые изучаются генетиками. Выбранные Менделем для анализа отдельные признаки гороха – гладкая и морщинистая форма семян, желтые и зеленые семядоли, серо‑коричневая и белая окраска семенной кожуры, выпуклая и с перетяжками форма боба, желтая и зеленая окраска зрелого боба, пазушное и верхушечное расположение цветков – дискретны, наследственны, альтернативны. Присутствие этих признаков служит основанием для выделения того или иного генотипа – эти признаки фенотипически обособляют его от других особей, а если рассматривать какую‑либо совокупность генотипов (в случае гороха – сорт, подвид), то по частоте присутствия этих признаков можно отличать и отдельные группы особей.
Рассмотрим некоторые из многих сотен известных наследственных дискретных вариаций строения тела у дрозофилы. Много признаков связано с формой крыла.

Рис. 3. Схема строения крыла дрозофилы
Точками показаны участки, наиболее подверженные изменчивости
У нормальной по строению мухи крыло всегда плоское. Среди изученных и генетически проанализированных особей встречались мухи‑мутанты с крыльями, изогнутыми в виде арки, закрученными кверху или книзу и с целым рядом других нарушений формы крыла. Часто встречаются мухи с вырезками на заднем крае крыла, с обрезанным задним краем крыла, с резко уменьшенными или вообще зачаточными крыльями. Есть мутантные мухи, вообще лишенные крыльев.
Множество наследственных вариаций у дрозофилы проявляется в особенностях жилкования крыльев: исчезновении отдельных продольных или поперечных жилок, разветвлении, утолщении и появлении добавочных жилок и т. д. (рис. 3).
Немало наследственных вариаций касается особенностей щетинок (макрохет), расположение которых является важным таксономическим признаком для большинства мелких насекомых. В результате мутаций отдельные щетинки исчезают вообще, другие удваиваются; возникают резко укороченные или вильчатые щетинки и т. п. Из других наследственных вариаций у дрозофил, бросающихся в глаза при внешнем осмотре, выделяют много признаков, связанных с окраской тела, видоизменением усиков, ног, брюшных сегментов, глаз.
Гены, ответственные за появление того или другого признака (в изученных нескольких сотнях мутаций), располагаются в определенных хромосомах, а внутри хромосомы – в определенных ее участках. Естественно, что признаки, закодированные в гомологичных хромосомах, чаще встречаются вместе (так называемые генетические группы сцепления) и признаки, которые встречаются в половых хромосомах, оказываются «сцепленными» с полом.
Проявление в фенотипе разных аллелей[16]одного гена делает возможным учет тех признаков фенотипа, которые можно называть фенами. Рассмотрим несколько других примеров фенов, наследственный характер которых изучен достаточно хорошо.
Красная и черная окраска двуточечной божьей коровки является парой хорошо заметных фенов. В то же время тщательные генетические исследования на одном из видов растительноядных божьих коровок показали, что здесь феном должен считаться угол наклона длинной оси пятен на покровных крыльях. Интересно, что генетический анализ тысяч особей не позволил выделить в фенотипе какие‑либо дискретные признаки, связанные с формой или величиной пятен у этого вида.
У млекопитающих множество фенов можно выделить на основании изучения скелета. Мелкие изменения формы отдельных костей, формы отростков, отверстий для кровеносных сосудов и нервов, наличие отдельных дополнительных так называемых брегматических костей в черепе – все это наследственно обусловленные дискретные признаки, которые можно назвать фенами.
Издавна для человека известны отклонения от нормального строения стопы или кисти, выражающиеся в уменьшении или увеличении числа пальцев. Появление шестого пальца руки связано с одной мутацией, сцепленной с полом. В некоторых семьях этот признак прослеживается на протяжении многих поколений.
Неизвестен наследственный характер расщепления четвертого или пятого пальца у дельфина белухи. Однако с большой степенью вероятности можно предположить, что это выражение особенностей генотипа. Исследования показали, что дальневосточные и северные популяции белухи отличаются друг от друга по частоте встречаемости этого фена.
Без сомнения, любой внимательный человек вспомнит множество примеров наследственных признаков, характерных для семей своих близких или родных. Такими признаками могут быть родинки на определенных местах тела, форма бровей, носа, губ. Выпяченная нижняя губа прослеживается в роду королевского дома Габсбургов на протяжении многих поколений.
Все приводимые выше примеры касались неметрических признаков, т. е. тех, которые нельзя измерить. Среди размерных признаков также можно выделить фены. Пример с карликовым и высоким растениями гороха, взятыми Менделем для анализа наследственности, хорошо иллюстрирует этот подход. В случае количественных признаков фенами могут быть такие признаки, которые образуют два или более дискретных, не перекрывающих друг друга варианта, например высота растений от 10 до 15 см, от 17 до 25 см, от 26 до 37 см.
В настоящее время накопилось множество данных, касающихся изучения чистых линий растений и животных. Среди этих данных немало примеров существования дискретных вариаций количественных признаков. Так, например, средний вес надпочечников в линии лабораторных мышей СЗН составляет 44,0±5,7 мг/100 г веса тела, а в линии мышей C57BL – только 16,0±1,2 мг/100 г.
Не следует думать, что фены могут быть только морфологическими признаками фенотипа. Фены могут касаться любых других признаков – и физиологических, и поведенческих. Один из интересных примеров физиологического фена – нечувствительность крыс к сильнейшему яду – антикоагулянту варфарину.
Этот фен был обнаружен в 50‑х годах после успешного истребления крыс в ряде стран Западной Европы в результате массового использования варфарина. Действительно, крысы в первое время практически исчезли в большинстве районов, где применялся варфарин. Но уже через несколько лет началось распространение крыс, нечувствительных к варфарину. Оказалось, что кое‑где в силу естественной генетической изменчивости в природных популяциях выжили отдельные группы особей, нечувствительные к варфарину. Подобная нечувствительность, как показали исследования, имеет разную генетическую природу и определяется по крайней мере действием трех генов. Однако эти три гена проявляются как один фен «нечувствительности к варфарину». В настоящее время крысы, нечувствительные к варфарину, широко распространены по всей Западной Европе. Это наглядный пример распространения нового адаптивного свойства под действием отбора в ответ на воздействие человека.
Описанные выше примеры касаются фенов животных. Но приведенные в предыдущей главе таблицы признаков, по которым вел свои исследования Н. И. Вавилов, восполняют этот пробел. Подавляющее большинство признаков у исследованных видов растений представляют собой типичные фены.
После такого предварительного обзора некоторых примеров фенов можно попытаться сформулировать более развернутое определение понятия «фен».
Ген и фен
Из приведенных примеров ясно, что можно называть феном: это дискретный, альтернативный признак, отражающий наследственные (генотипические) особенности особи.
Чтобы быть точным и последовательным, следует прежде всего сказать о самом понятии «признак». Как это бывает, самые простые, часто используемые понятия точно определить довольно сложно. В логике признаком называют «все то, в чем предметы, явления сходны друг с другом или в чем отличаются друг от друга…»[17]. Главное отличие фена от остальных признаков – его диагностическая ценность: по присутствию того или иного фена можно составить представление о генотипе.
Однако положение с определением фена сложнее, чем может показаться на первый взгляд. В общей форме совершенно ясно, что любой признак имеет в той или иной степени наследственную обусловленность: нет таких особенностей фенотипа, возможность появления которых не была бы запрограммирована в генотипе. Таким образом, наследственно обусловлены не только альтернативные, дискретные признаки, но и вес, и длина, и пропорции отдельных частей тела.
Однако совершенно ясно, что наследственная обусловленность размера тела чем‑то существенным отличается от наследственной обусловленности цвета глаз. Генетическая обусловленность размера тела свиньи включает возможность формирования при хороших условиях борова весом в 200 кг, а при плохих условиях – весом всего в 50 кг. Этот диапазон возможных вариантов при одном и том же генотипе показывает широту нормы реакции. В ее пределах изменчивость признака контролируется условиями развития. Можно сказать, что наследуются не признаки, а норма реакции – способность к формированию тех или иных признаков при определенных условиях развития.
Тут мы подходим к интереснейшей и малоразработанной проблеме взаимоотношения гена и признака, к самой большой Terra incognita в современной биологии – проблеме наследственного осуществления.
Конечно, сложность реализации наследственной информации, работы генетического кода настолько велика, что трудно было бы ожидать однозначных и исключительно постоянных соотношений между геном и феном. Иногда оказывается, что под внешне однородным фенотипом скрывается действие разных генов. Известно, например, что фенотипическое проявление мутации black и мутации ebony у дрозофилы практически одинаково – появляются мухи с черным телом. Только генетический анализ позволяет различить эти мутации, локализованные в разных хромосомах. Несколько разных мутаций могут вызывать бесхвостость домовых мышей. При внешнем анализе фенотипа исследователь сможет отметить лишь один фен бесхвостости. Эту возможность маскировки разных генов одним и тем же феном всегда нужно учитывать.
Принципы полимерии (на каждый признак влияют несколько генов) и плейотропии (каждый ген влияет на несколько признаков) были известны с начала века. Это – феноменология всего процесса, наблюдаемый конечный результат. На рис. 4 схематически изображено действие этих принципов в развитии особи. Таких схем даже для не особенно полно изученных генетически видов животных и растений уже полстолетия тому назад можно было составить довольно много, а в наше время – сотни.
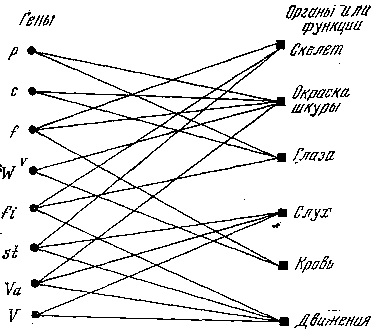
Рис. 4. Схема соотношения генов и признаков у домовой мыши
Р – красный цвет, с – альбинизм, f – согнутый хвост, Wv – доминантная пятнистость, fi – беспокойное поведение, st – трясучесть, Va – переваливающаяся походка, V – вальсирующее передвижение
Подробно рассмотрим только один пример плейотропии, связанный с геном карликовости у мышей. Это рецессивный ген, определяющий развитие животных, в два‑три раза меньше нормальных размеров; он возник в одном из питомников белых мышей в 1929 г. Еще до остановки роста мышей‑карликов можно отличить по целому ряду признаков: тупым мордочкам, коротким ушам и хвостикам, вялости, робости, чувствительности к колебаниям температуры и другим признакам. Продолжительность жизни карликовых мышей короче: и самцы и самки стерильны.
Этот яркий пример плейотропии в действии гена интересен одной особенностью. Оказалось, что карликовых мышей можно превратить в нормальных, если хирургически вживлять в их тело (под кожу) кусочки гипофиза крысы. (Гипофиз – небольшая железа внутренней секреции, расположенная в основании мозга и регулирующая выделение практически всех гормонов.) После ряда таких, операций карликовые животные преображались: они достигали нормальных размеров и начинали вести себя как нормальные мыши (правда, самки оставались стерильными).
Этот пример показывает, как наследственно определенные признаки могут резко измениться под влиянием среды, внешних условий. Значит, если выразиться точнее, не аллель карликовости ведет к стерильности, ранней остановке роста, стойкости к голоду и повышенной чувствительности к холоду, а действие аллеля карликовости ведет к такому изменению нормы реакции, при котором в обычных условиях развивается стерильность, рано прекращается рост, повышается стойкость к голоду и чувствительность к изменениям температуры.
Рассмотренный пример показывает также, что действие внешних по отношению к генотипу агентов способно замаскировать то или иное проявление гена. Эту возможную маскировку фенов, так же как и противоположный случай – возникновение псевдофенов – дискретных признаков в результате каких‑то внешних вмешательств в развитие (например, травматические изменения), нужно постоянно иметь в виду. Такая всегда существующая неопределенность в выделении фенов – та цена, которую исследователю приходится платить природе при отказе от изучения каждого признака генетически, с применением методов скрещиваний в чреде поколений. Однако преувеличивать значение этой неопределенности для фенетического анализа не следует. Далее будет рассказано о приемах, позволяющих ослабить ее влияние при анализе конкретных данных.
Пока нет сколько‑нибудь надежного сравнительного материала, позволяющего количественно оценить маскировку разных генов проявлением одного и того же фена. Однако, поскольку число признаков фенотипа практически бесконечно, а число генов у каждого вида имеет конечный характер, в будущем, при развитии фенетических методов анализа фенотипа, всегда можно найти фены, маркирующие ген либо непосредственно и однозначно, либо посредством сочетания. Но это – дело будущего.
Важнейшее значение для понимания природы фенов имеют работы английского генетика Г. Грюнеберга, проведенные в 50–60‑х годах. Ученый сравнивал разные линии мышей (максимально однородные генетически в результате близкородственного разведения) по встречаемости мельчайших вариантов в строении скелета. Этими вариантами было развитие какого‑либо отростка на костях, прохождение группы кровеносных сосудов в данном месте скелета через одно большое или несколько малых отверстий, расположение определенных отверстий для прохождения кровеносных сосудов и нервных стволов, различные аномалии зубной системы и т. п.
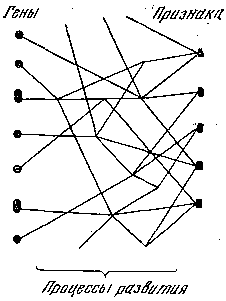
Рис. 5. Уточненная схема взаимоотношений гена и фена
Оказалось, что все варианты относились к числу пороговых: как только в процессе эмбрионального развития тот или иной зачаток будущей структуры достигал определенной величины, осуществлялось формирование будущей структуры – возникал отросток на кости, большое отверстие разделялось перегородкой надвое и т. п. Если же зачаток структуры в эмбриогенезе не достигал этой величины, подобные признаки не появлялись у взрослого организма. Хотя на величину зачатка оказывали влияние многочисленные взаимодействующие гены и внешние факторы, решающей причиной в конце концов приходилось считать генетическую. Изученные линии мышей хорошо различались по частотам таких фенов, и эти различия передавались по наследству.
Теперь, после более подробного рассмотрения связи гена и фена становится ясно, что изображенная на рис. 4 схема соотношения между генами и признаками нуждается в уточнении. Эти соотношения оказываются более сложными: плейотропия и полимерия представляют собой не свойство самих генов, а являются итогом различных реакций (от простейших биохимических до сложнейших морфогенетических), управляемых генами (рис. 5). Фены представляют собой видимые, улавливаемые наблюдателем звенья таких реакций.
Обычно в качестве фенов описываются конечные звенья морфогенетических цепей – «конечные продукты действия генов», по выражению австрийских исследователей Бергеля и Наса (1976). Однако дискретными признаками характеризуются и отдельные этапы в процессе индивидуального развития организма. Такой подход приложим к фенам у растений, как показано недавно работами по фенетике растений дагестанского исследователя М. Магомедмирзаева (1976–1977), который в изменчивости травянистых растений и деревьев выделяет элементарные, дискретные процессы, так называемые кванты морфогенеза.
После всего сказанного выше можно дать более полную формулировку понятия «фен». Феном называются любые дискретные альтернативные вариации признаков и свойств живых организмов, которые на всем имеющемся материале (обязательно многочисленном) далее неподразделимы без потери качества. Фены всегда отражают определенные черты генетической конституции данной особи, а своей частотой – генетическую структуру популяции и других (как более, так и менее крупных) групп особей данного вида.
Как найти фены?
В генетике для анализа того или иного признака достаточно скрестить пару особей и по характеру распределения признаков в потомстве (на протяжении ряда поколений) точно определить особенности любого признака или свойства. Для фенетического анализа необходимо значительное число особей. Без них исследователь не сможет составить достаточно полное представление о существующих в природе дискретных вариациях, среди которых и приходится искать фены.
Опыт, накопленный при исследовании фенов в разных группах животных и растений, позволяет предложить следующие пути выделения фенов на материале природных популяций.
1. Сопоставление наблюдаемой изменчивости по отдельным комплексам признаков или свойствам (окраске, рисунку, форме отдельных частей тела, вариантам строения отдельных органов и систем и т. п.).
2. Выделение признаков, подверженных возрастным и половым изменениям. Для первичного анализа они сложны, хотя среди них тоже можно найти фены, но это – более специальная задача. Обычно такие признаки не включаются в фенетическое исследование на животных, хотя, как упоминалось выше, могут быть с большим успехом использованы у растений.
3. Выделение в общей изменчивости признаков и свойств дискретных характеристик, их анализ на дальнейшую делимость (дробимость, подразделяемость).
Предположим, что изучаются такие особенности окраски, как рисунок и цвет. Дискретность в изменчивости нужно искать именно в цветовой гамме, а не в микроскопически определяемых характеристиках пигмента или микроструктуре поверхности органа, вызывающего цветовые эффекты (как у чешуекрылых). Равным образом было бы неправильно свести рисунок окраски (полосы, пятна на отдельных участках тела и т. п.) к изучению, например, гистологии кожного покрова. И в том и в другом случае исследователь потеряет качество изучаемой группы признаков и перейдет к изучению совсем другой группы признаков и свойств (среди которых тоже можно искать качественные альтернативные дискретные признаки).
4. Анализ данных по генетике филогенетически близких генетически изучаемых форм. Чтобы проанализировать окраску рыжей полевки, нужно познакомиться с работами по генетике окраски других грызунов (домовая мышь, крысы и др.). Зная закон гомологических рядов в наследственной изменчивости, можно считать, что общие закономерности окраски млекопитающих будут в чем‑то существенном общими. За цвет покровов у млекопитающих несут ответственность шесть основных, сходных у разных видов млекопитающих генов, а за рисунок – еще несколько таких же генов. Конечно, у каждого вида несколько основных генов могут дополняться другими, их действие может в разной степени модифицироваться, но сам факт наличия гомологичных генетических основ позволяет уверенно вести фенетический анализ окраски практически всех млекопитающих.
5. Анализ косвенных данных о характере наследования отдельных признаков у данного вида (выражение признаков у родителей и потомков и т. п.).
6. Проверка правильности выделения фенов на природном материале. Обычно правильно выделенные фены на достаточно разнообразном материале из природных популяций обязательно покажут или определенные тенденции в распространении их в пределах ареала вида, или изменение во времени (на палеонтологическом материале).
Теперь рассмотрим описанный порядок выделения фенов на конкретных примерах.
Пример I. Цель исследования – анализ внутривидовой изменчивости окраски (цвета) покрова прыткой ящерицы для выяснения особенностей процесса микроэволюции на уровне отдельной популяции, групп популяций и всего вида в целом. Таким образом, представляли интерес признаки от самого мелкого до самого крупного масштаба.
1. Изучение вариантов окраски по цвету покровов из нескольких природных популяций показало присутствие широкого спектра серо‑зеленых и желто‑коричневых тонов в окраске спины, бело‑голубовато‑зеленых в окраске брюха, бело‑желтых в окраске горла.
2. Сопоставление окраски молодых и взрослых животных показало, что существуют значительные возрастные изменения окраски – окраска спины молодых однородно коричнево‑серая. Кроме того, окраска брюха и горла также изменяется с возрастом. Поэтому для дальнейшего анализа использовались лишь половозрелые особи, легко выделяемые по размерам тела.
Исследование окраски самцов и самок показало сложную картину: в одних популяциях половой диморфизм и окраске спины выражался резко (самцы зеленые, самки коричневые), в других – и среди самцов и среди самок встречались коричневые и зеленые особи. Сложным оказался и признак окраски горла: как правило, самцы имели более ярко окрашенное горло (ярко‑желтое или даже зеленое), но нередко встречались и самки с желтым горлом. Окраска брюха у самцов более зеленая, чем у самок, но варьировала и у того и у другого пола весьма значительно.
3. Очень широкий спектр цветовых вариаций включал первоначально восемь цветовых оттенков для окраски спины (темно‑зеленый, ярко‑зеленый, светло‑зеленый, желтый, светло‑коричневый, красноватый, коричневый, темно‑коричневый) и по два‑три оттенка окраски брюха и горла. Для проверки правильности отнесения той или иной особи к определенной группе окраски были составлены специальные цветовые таблицы. Однако практика применения таблиц показала субъективность восприятия разными наблюдателями выделенной первоначально цветовой гаммы. Безусловно совпадающим у всех исследователей оказалось лишь выделение зеленого и коричневого цветов в окраске спины (впоследствии в качестве очень редких вариантов были найдены «красные» – кирпичного цвета – и черные особи). В качестве дискретных признаков были выделены зеленые и коричневые тона в окраске спины, и пришлось полностью отказаться от точного учета окраски брюха и горла.
4. Имеющиеся в литературе данные по наследованию окраски у прыткой ящерицы касались менделевского характера наследования кирпично‑красной окраски спины. Данные по генетике окраски ящериц другого рода (американских Sceloporus graciosus) показали высокую наследственную обусловленность песчано‑красной окраски спины.
5. Косвенные данные по наследованию окраски в природе и эксперименте практически отсутствовали из‑за значительного возрастного полиморфизма в окраске и несравнимости окраски взрослых и неполовозрелых особей. Известны наблюдения единичных черноокрашенных и молодых и взрослых особей в одних и тех же местообитаниях (это позволило предполагать их близкое родство).
6. Анализ частоты встречаемости зеленого и коричневого цвета в окраске спины у самцов и самок показал в целом определенную закономерность: на севере и самцы и самки были окрашены в коричневый цвет, на юге практически все самцы имели зеленую окраску, тогда как среди самок встречались и коричневые и зеленые особи. Иными словами, и для самцов и для самок было установлено закономерное изменение частоты встречаемости зеленой и коричневой окраски с юга на север, причем характер этой закономерности у самцов и самок отличался. Таким образом, фены окраски показали различие в масштабе крупных частей ареала вида. При анализе же внутри отдельных популяций эти признаки не дали каких‑либо определенных результатов – их распределение было хаотично.
Пример II. Этот пример касается анализа изменчивости швов черепа гренландского тюленя. Была поставлена задача найти доказательства популяционной самостоятельности трех группировок этого вида в Северной Атлантике. Имевшийся материал включал черепа животных разного возраста (от 2 до 36 лет), добытых во всех трех предполагаемых популяциях.
1. Сопоставление всей наблюдаемой изменчивости показало, что швы бывают полностью заросшими, без каких‑либо следов соединения костей, бывают хорошо заметными и разной формы (рис. 6). Кроме того, встречались черепа, у которых на правой половине швы были хорошо развиты, а на левой отсутствовали.
2. Сравнение черепов разновозрастных особей показало, что заращение швов не имеет прямой связи с возрастом: у самых старых животных были найдены хорошо выраженные швы, тогда как у молодых в некоторых случаях швы зарастали полностью. Половой диморфизм в развитии швов не обнаружен. Все это позволило оставить для дальнейшего анализа все первоначально выделенные признаки присутствия или отсутствия швов хотя бы на одной стороне и их формы.

Рис. 6. Разная форма швов в черепе млекопитающих
3. Присутствие швов оказалось дискретным признаком: не было найдено животных с их частичным присутствием.
4. Анализ данных по генетике показал, что некоторые линии домовых мышей различались по степени зарастания швов к моменту достижения половой зрелости (у всех мелких млекопитающих с возрастом швы зарастают). Из антропологии известны случаи наследственного сохранения несросшимися швов черепа в течение всей жизни. В литературе имеются данные о нахождении незаросших швов черепа у старых особей самых разных видов млекопитающих, в том числе у многих хищных – отряда, филогенетически близкого к ластоногим.
5. Данные по сравнению характера швов черепа у родителей и потомков из природных популяций отсутствовали.
6. Анализ частоты встречаемости разного типа швов в трех популяциях гренландского тюленя показал существование различий между популяцией о‑ва Ньюфаундленд, с одной стороны, и Гренландского и Белого морей – с другой. Эти различия выявлены при сравнении встречаемости незаросших швов, а не форм швов. Фен присутствия швов оказался маркером, позволяющим подтвердить самостоятельность одной из изученных популяций.
Кратко охарактеризованный путь выделения фенов на зоологическом материале применим и для работы с растениями. При выделении фенов важно обратить внимание на их достаточную многочисленность.
Для решения каких‑то сравнительно простых вопросов популяционного исследования иногда бывает достаточно одного фена. Например, в тех случаях, когда фен в одной популяции присутствует в высокой концентрации, а в другой – отсутствует или присутствует в ничтожных концентрациях. В этих случаях уже можно сделать обоснованное предположение о существовании границ между исследованными совокупностями особей.
Однако опыт работы Н. И. Вавилова с разными видами растений, наш опыт работы с ящерицами, анализ наиболее успешных случаев использования фенетических методов на других видах показывает, что для надежной и полной характеристики всего диапазона пространственной изменчивости (от выделения отдельных семей до выделения подвидов) надо учитывать многие десятки (а то и несколько сотен) признаков. Тогда есть полная уверенность, что любая задача популяционного анализа будет успешно решена.
Не надо бояться потерять много времени на выделение фенов. Дело в том, что все сказанное относится именно к исследованиям какой‑либо группы, ранее не изучавшейся с целью выделения признаков, которые могут быть приняты как фены. В группах, где проведены такие исследования, ситуация складывается скорее обратная. Если по какому‑либо виду уже собран фенетический материал (другими исследователями, в других районах и в другое время), то каждая последующая работа оказывается способной привести ко все более интересным и глубоким выводам. Возникает полная аналогия с выкладыванием картины из мозаики: после добавления одного‑двух элементов прежде казавшиеся рассыпанными в беспорядке частицы складываются в общую осмысленную картину, проявляются сложные узоры и рисунки.
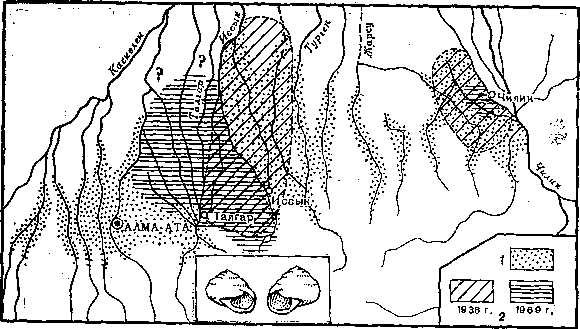
Рис. 7. Распространение право‑ и левозакрученных форм наземного моллюска и изменение ареала левозакрученных форм в Заилийском Алатау на протяжении 30 лет (с 1936 по 1969 г.)
1 – правозакрученные моллюски, 2 – левозакрученные
Приведем только один пример, касающийся исследований фена – левозакрученности раковин у наземных легочных моллюсков (Fruticicola lantzi) в предгорьях Заилийского Алатау. Левозакручениость – типичный фен, имеющий у всех изученных других видов моллюсков моногенную основу. Распространенность этого фена в популяциях Средней Азии первоначально изучена в конце 30‑х годов, повторно – в конце 60‑годов. Оказалось, что в одной группе популяций территория, занятая особями с этим феном, во много раз сократилась, а в другой – сдвинулась на несколько десятков километров (рис. 7). Каждое новое исследование этого интересного природного эксперимента будет все более простым и в то же время информативным, позволяя выяснять динамику генетического состава популяций.
Масштаб фенов
Фены могут быть разного масштаба. Есть фены, своим присутствием маркирующие отдельные особи и самые мелкие из возможных естественных группировок – потомство одной пары родительских особей. Такими «семейными» фенами были и выпяченная губа Габсбургов, и ненормальный палец на руке у герцогов Шрюсбери.
На сравнительно небольшой площади в несколько квадратных километров было просмотрено несколько тысяч особей прыткой ящерицы. При этом отмечались все встречавшиеся особенности окраски (рисунка), а также форма и расположение щитков на голове и других участках тела. Всего учитывалось в полевых условиях несколько десятков признаков, среди которых, как мы предполагали, могли быть и фены. Среди значительного числа просмотренных особей только у трех был найден наданальный щиток в виде пятигранника особой формы. И все эти три особи были пойманы на расстоянии нескольких метров друг от друга. Единственное возможное объяснение этого – предположение, что наданальный щиток оказался признаком семейного масштаба.
По сочетанию и форме отдельных пятен на голове, горле и брюхе зоологи научились «узнавать в лицо» отдельных ласок. Недавно выяснилось, что по расположению вибрисс на морде можно даже издалека узнавать львов в природе.
Один из замечательных примеров семейных фенов описан у кашалотов. Среди более чем 5 тыс. просмотренных особей были добыты две самки с совершенно одинаковым рисунком, ни до того, ни после того не встреченным, – резкая поперечная белая полоса на боку тела (рис. 8). Эти самки были добыты вместе, из одной группы. У одной из самок оказался эмбрион с точно таким же характером окраски. Эта замечательная находка сразу же дала возможность решить задачу по биологии кашалотов, до того времени остававшуюся предметом жарких споров среди специалистов. Она показала, что самки, входящие в гарем – группу из 10–30 самок с двумя‑тремя крупными самцами, могут быть родственниками, а весь гарем – семейным объединением. Находка же матери и детеныша с одинаковым редким рисунком, указывающим на наследственную природу этого признака, позволила говорить о нем как о настоящем фене.
Итак, самые мелкие, индивидуально специфические, редкие вариации каких‑либо признаков дают в руки исследователя фены семейного порядка.
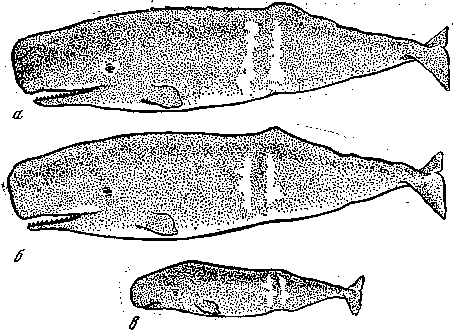
Рис. 8. Пример фенов окраски семейного масштаба у кашалотов, добытых из одной группы (гарема)
а, б – две взрослые самки, в – предродовой эмбрион
В любом достаточно обширном исследовании можно обнаружить и фены большего масштаба. У животных это будут фены, маркирующие демы, – группы, состоящие из нескольких семейных группировок (или еще более крупные и сложные), занимающие определенное пространство внутри популяции, сохраняющие относительную самостоятельность на протяжении жизни немногих поколений. Такие группировки были найдены у прыткой ящерицы на Алтае: несколько из учитываемых фенов показали различную концентрацию в разных долинках между соседними холмами. В других случаях исследователи находили статистически достоверные различия по распространенности разных фенов для групп особей внутри популяций у ряда мышевидных грызунов и других видов животных.
В следующей главе мы проанализируем примеры выделения отдельных популяций и групп популяций по частоте распространенности разных фенов. Крайние случаи такого рода – фены видового и более крупного масштаба.
Можно заметить, что редкие фены позволяют, с одной стороны, различать какие‑то очень мелкие группы особей внутри популяции, а с другой стороны – достаточно крупные группы. Например, фен совершенно черной окраски тела (полный меланизм) встречен у прыткой ящерицы на Калбинском Алтае и в окрестностях Барнаула. Этот фен одновременно позволяет различать демы и всю большую группу алтайских популяций вида.
Фены, встречающиеся со средней или высокой концентрацией, видимо, пригодны лишь для сравнения достаточно большого числа групп особей.
Итак, фены бывают разного масштаба – от индивидуальных и семейных до видовых. Возможно, они объединяют и несколько видов: в палеонтологическом материале, где фены могут характеризовать отдельные небольшие ветви древа жизни.
Дата добавления: 2016-01-26; просмотров: 2721;