Специальные гены в эволюции
Адаптируемость – это способность вида поднять уровень своей абсолютной приспособленности в неоптимальных условиях среды. Что же такое абсолютная приспособленность вида и как определить ее уровень? Пока не существует строгого определения этих понятий. По‑видимому, лучше всего представление об абсолютной приспособленности передает величина биомассы (или количество особей), производимой видом за единицу времени. Очевидно, что эта характеристика прямо связана с биомассой вида (или с численностью его особей).
Для того чтобы проследить за динамикой уровня абсолютной приспособленности вида, мы должны иметь возможность наблюдать за ним в течение многих десятков (и даже сотен) поколений. Фактически нам следовало бы наблюдать всего за двумя характеристиками вида – географическим ареалом и числом особей в его популяциях. Обычно плотность популяций максимальна в центре ареала, откуда некоторый «избыток» особей перебрасывается миграционным потоком на периферию, где гибнет под влиянием неблагоприятных факторов. Если мощность производства биомассы в центре ареала ослабевает, то падает средняя плотность популяций и сокращается площадь видового ареала. Конечно, при ежегодных наблюдениях мы бы заметили довольно резкие колебания обеих характеристик, но если бы их средние многолетние значения достоверно не снижались, то можно было бы сказать, что данный вид неплохо приспособлен к среде своего обитания.
Теперь проведем один мысленный эксперимент: истребим всех особей вида, кроме одной оплодотворенной самки. Поместим ее в искусственно созданные идеальные условия, многократно размножим потомков и выпустим их обратно в места обитания предков. Едва ли кто усомнится в исходе этого предприятия: вероятнее всего, вид вскоре восстановит свой прежний ареал и численность. В успехе подобного эксперимента нас убеждают многочисленные случаи восстановления совсем было истребленных видов (выхухоль, бобр, зубр, бизон и т. д.). Кроме того, о силе жизни, заключенной в считанных штуках особей, свидетельствуют примеры успешной интродукции вида (иногда совсем нежелательной) в новые места обитания. Кролики в Австралии, лошади в Америке, колорадский жук в Европе, рапана в Черном море и масса других подобных случаев. Везде мы видим, что основные характеристики, определяющие абсолютную приспособленность вида, заключены в геноме типичной, т. е. фактически любой особи.
Пожалуй, наиболее ярко об этом свидетельствует «успех» канадской элодеи, впервые обнаруженной в Ирландии в 1836 г. У себя на родине эта элодея представлена мужскими и женскими растениями, т. е. ей вполне свойственно половое размножение, кроме того, каждое растение (точнее, любой фрагмент его стебля с несколькими мутовками) способно к неограниченному росту и ветвлению. Завезенный в Европу женский экземпляр, размножаясь только вегетативно, породил столько потомков, что их биомасса вскоре стала мешать судоходству в каналах Англии. В настоящее время канадская элодея уже достигла рек и озер Восточной Сибири. Сила этого единственного генотипа отражена в названии, данному растению в Европе, – «водяная чума». Правда, сейчас его мощь явно ослабела, вероятно, стал сказываться мутационный груз, для сброса которого нужен половой процесс.
По‑видимому, главное, что несет в себе типичная особь вида, заключается в скорости, с которой организм в конкретных условиях среды воспринимает извне вещество и энергию, преобразуя их в биомассу потомков. Что же определяет эту скорость? Очевидно, только мощность рабочих структур типичной особи. Напомним, что в гл. 2 мы определили такую мощность, как объем работы, выполняемой структурой за единицу времени в режиме максимальной нагрузки. При ухудшении среды мощность какой‑то рабочей структуры может оказаться недостаточной, поэтому численность вида и его ареал сократятся, и восстановить их будет возможно, лишь усилив мощность лимитирующей структуры, ставшей фактически одной из главных компонент приспособленности. Выходит, абсолютную приспособленность вида можно было бы выразить в единицах мощности.
В данной ситуации мутации, повышающие мощность лимитирующей структуры, очевидно становятся благоприятными, так как они повышают уровень относительной приспособленности их носителей. Поэтому коэффициент отбора (s ) в пользу этих мутаций должен быть близок к относительному приросту мощности лимитирующей структуры, во всяком случае, он должен быть ему пропорционален, т. е.
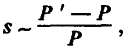
(5.3)
где Р ' – мощность структуры у особей – носителей благоприятной мутации; Р – мощность той же структуры у средней особи популяции. Фактически это означает, что фиксация каждой благоприятной мутации увеличивает мощность лимитирующей структуры в некоторое число раз (на какой‑то процент). При продолжающемся ухудшении среды продолжается и процесс накопления благоприятных мутаций. В итоге мощность лимитирующей структуры нарастает вдоль филетического маршрута как ряд:
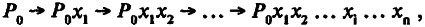
(5.4)
где Р 0 – мощность структуры до начала действия движущего отбора; x i – число раз, в которое увеличивается мощность структуры в ходе фиксации i ‑й мутации. Это как раз те «незначительные изменения», аккумуляция которых, по выражению Дарвина, ведет к созданию органа «любой мыслимой степени совершенства».
Заметим, что логарифм мощности структуры (lnP ) после фиксации и благоприятных мутаций определяется равенством
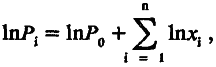
(5.5)
где стоящий под знаком суммы lnxi представляет собой вклад i ‑й благоприятной мутации в логарифм мощности рабочей структуры; для удобства дальнейшего изложения этот вклад будем обозначать аi'. Если единицей измерения мощности рабочей структуры выбрать ее исходную величину Р0, то (учитывая, что lnl = 0) уравнение (5.5) можно представить в более простой форме:
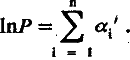
(5.6)
Напомним, что аi ' – вклад i ‑й мутации, зафиксированной отбором, направленным на усиление лимитирующей структуры.
Теперь введем одно определение: локусы, по которым шло замещение новых аллелей, повышающих мощность структуры в ее историческом развитии, назовем специальными генами структуры. Вполне очевидно, что некоторые из этих генов могут повторно мутировать, порождая новые аллели, усиливающие лимитирующую функцию. Таким образом, среди n благоприятных мутаций, зафиксированных движущим отбором, далеко не все связаны с увеличением числа специальных генов N.
Представим себе, что изменение среды приняло противоположное направление. В новых условиях мощность структуры, развитой за предыдущий период, может стать избыточной, поэтому ничто не будет препятствовать накоплению мутаций, ослабляющих экспрессию ее специальных генов. Более того, энергия и вещество, которые ранее расходовались на поддержание высокой мощности этой структуры, теперь могут быть направлены на создание дополнительной биомассы, т. е. на повышение абсолютной приспособленности вида. Таким образом, ослабление рабочей структуры в данном случае представляет собой адаптивный ответ вида на «требование» изменившейся среды. Через некоторое время новая среда может снова потребовать усиления функции той же структуры, и снова благоприятными станут мутации, усиливающие экспрессию ее специальных генов.
Таким образом, для вида, предки которого прошли через многократные изменения требований среды к интенсивности функции определенной рабочей структуры, логарифм ее мощности можно представить в виде суммы вкладов N специальных генов, где вклад каждого такого гена (его экспрессия ) (а ) может изменяться от нуля до какого‑то максимального значения, т. е.
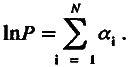
(5.7)
Это выражение удобнее представить в несколько иной форме:
lnP = Na, (5.8)
где а – уровень средней экспрессии специального гена, т. е. его средний вклад в логарифм мощности рабочей структуры. У разных видов а может варьировать в очень широком диапазоне: от нуля, когда все специальные гены «выключены», до величины, соответствующей уровню их максимальной активности. Ясно, что уровень средней экспрессии специального гена рабочей структуры зависит от предыстории вида, от того, как долго среда «требовала» от предков усиления мощности структуры и как долго была к ней безразлична или даже «ждала» ослабления.
Пусть мы имеем дело с совокупностью родственных видов одной филетической группы. Рассмотрим у них какую‑нибудь гомологичную структуру с одной и той же функцией. У каждого вида (в соответствии с (5.8)) логарифм мощности структуры равен произведению Na. Теперь перейдем к среднему (для видов филетической группы) логарифму мощности структуры (<lnP >), т. е. от каждого вида возьмем значение логарифма мощности структуры ее типичной особи, полученные величины сложим, а их сумму разделим на число видов в группе. В теории вероятностей доказывается, что среднее значение произведения независимо варьирующих сомножителей равно произведению их средних значений. Следовательно, средневидовой логарифм мощности структуры будет равен средневидовому уровню экспрессии ее специальных генов (<а >), умноженному на среднее число этих генов у одного вида (<N >), т. е.
<lnP > = <N > <а >. (5.9)
Если линии филетической группы уже неоднократно сталкивались с изменениями требований среды к мощности какой‑то рабочей структуры, то средневидовой уровень экспрессии ее специальных генов можно считать в какой‑то степени стабилизированным. В этом случае соотношение (5.9) упрощается до
<lnP > ~ <N >. (5.10)
Число специальных генов, определяющих развитие структуры, можно принять за меру ее генетической сложности, поэтому соотношение (5.10) означает, что средневидовая мощность структуры, измеренная логарифмической шкалой, пропорциональна ее средневидовой генетической сложности.
Дата добавления: 2016-01-26; просмотров: 896;