Формирование видовых распределений
Построим простейшую модель видовых распределений. Будем считать, что для экологической дифференциации видов их особи должны отличаться по размерам на 28 %, соответственно натуральный логарифм их размеров – приблизительно на 0,25. Разобьем горизонтальную ось, вдоль которой изменяется логарифм линейных размеров, на интервалы шириной 0,25. Пусть вид‑основатель филетической группы по размеру своих особей находился в интервале х0. Через единичный отрезок времени (через один шаг модели) он произведет два дочерних вида, которые попадут в соседние интервалы х ‑1, и х 1. Отметим, что интервал х0 остается занятым материнским видом. В следующий временной отрезок каждый из трех видов приступит к «завоеванию» своих соседних интервалов. Продолжая такой процесс, мы вскоре получим довольно гладкое симметричное (биномиальное) распределение, очень близкое к нормальному. С каждым шагом оно расширяется, его дисперсия увеличивается пропорционально возрасту таксона, а сигма следует за квадратным корнем из этого возраста. Однако центр тяжести распределения (его среднее значение) должен оставаться на месте, точно соответствуя величине признака у вида‑основателя филетической группы – х0 (рис. 18, а).
Ясно, что такая модель не объясняет правила Копа. Начнем ее усложнять. Быть может, мы получим искомый сдвиг распределения вправо, если зафиксируем его левую границу, т. е. примем, что вид с величиной тела меньше некоторого критического значения х* просто не может существовать. Подобное соображение выглядит вполне реалистичным. Например, для очень мелких теплокровных животных, таких как колибри у птиц и землеройки у млекопитающих, дальнейшее уменьшение размеров создало бы серьезные проблемы с поддержанием температуры тела. По‑видимому, колибри, питающиеся очень калорийным нектаром, уже достигли левой границы распределения птиц. Кстати, в покое они не в состоянии поддерживать постоянную температуру.
Пусть критическое значение признака соответствует интервалу x‑8 . Отсюда следует, что вид, занимающий этот самый левый интервал, может дать дочерний вид только с увеличенным размером особей, заселяя интервал х‑7 . С ходом времени мы получим распределение, которое будет весьма походить на предыдущее, отличаясь от него практически только отсутствием части, расположенной слева от «поглощающего экрана» (рис. 18, б ). Естественно, с ходом времени среднее значение такого урезанного распределения будет смещаться вправо, а незначительным сдвигом его вершины (моды) можно пренебречь. Однако тщательный анализ реальных видовых распределений (подобных представленным на рис. 17) обнаруживает четкое смещение их вершины вправо, т. е. в сторону увеличения признака.
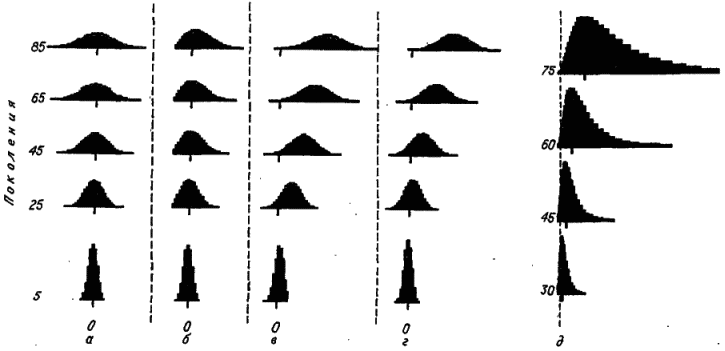
Рис. 18. Моделирование динамики видовых распределений по логарифму величины количественного признака. а – вероятность изменения признака в обоих направлениях равна и не зависит от его величины; б – те же условия, но признак не может принимать значения меньше, чем x‑8; в – ортоселекция (вероятность увеличения признака в 2 раза выше вероятности его уменьшения); г – рост адаптируемости (вероятность изменения признака в обоих направлениях равна и возрастает на 0,01 с переходом в соседний интервал справа); д – те же условия, что и в случае г (признак измеряется арифметической шкалой).
Есть два пути получить динамику распределений, близкую к реальной. Первый состоит во введении асимметрии в процесс заселения смежных интервалов, второй – в ускорении заселения обоих соседних интервалов (вполне симметричного) по мере увеличения размеров особей. Первый путь – это ортоселекция, когда дочерний вид заселяет быстрее интервал справа, чем интервал слева. Если мы предположим, что вероятность заселения правого интервала в 2 раза выше, чем левого, то получим картину, представленную на рис. 18,в . Видно, что распределение быстро расползается вправо и в ту же сторону сдвигается его вершина.
Обычно современные приверженцы синтетической теории эволюции считают, что за эти макроэволюционные тенденции ответственна именно ортоселекция. Их соображения выглядят примерно так. Увеличенные размеры тела дают индивидам очевидные преимущества: их бег становится быстрее и экономичнее, они лучше защищены от хищников и, наконец, они, обладая большей мышечной силой, имеют преимущества в драках с конкурентами за спаривание. Казалось бы все логично, хотя несколько антропоморфно. Действительно, людям свойственно воспевать крупных мужчин, их, так сказать, боевые качества. Попробуем посмотреть на это с иной точки зрения.
Увеличенная масса тела затрудняет бег по пересеченной местности, к тому же крупный индивид требует для поддержания жизни больше пищи. Кроме того, с увеличением массы тела, безусловно, падает скорость реакции на внешние раздражители – весьма немаловажное обстоятельство для выживания. Едва ли существует положительная корреляция между размерами тела охотника и количеством приносимой им добычи – основы благополучия первобытного человека. Наконец, увеличенные размеры означают некоторое удлинение онтогенеза и соответственно снижение шансов достичь взрослой стадии. Таким образом, без точных оценок всех плюсов и минусов мы не в состоянии найти простую связь между размерами особи и ее приспособленностью. Единственной твердо установленной закономерностью, связывающей размеры тела животных с приспособленностью, является правило Бергмана, согласно которому среднепопуляционная масса тела у теплокровных животных одного вида увеличивается, с понижением среднегодовой температуры. Этому правилу подчиняется большинство видов птиц и млекопитающих, верно оно и для людей.
Рассмотрим подробнее еще один путь, ведущий к результату, очень сходному с ожидаемым при ортоселекции. Пусть вероятность перехода дочерних видов в оба соседних интервала растет в прямой пропорции с номером интервала, в котором находился материнский вид. Заметим, что тем самым мы ввели предположение о существовании прямой зависимости между средним размером тела особи и адаптируемостью вида. На рис. 18, г представлена динамика таких распределений для случая, когда адаптируемость вида при переходе в соседний интервал справа возрастает на 0,01. Мы пришли к результату, фактически не отличимому от предыдущего: распределение расширяется вправо при переносе вправо и модального класса. Перейдя от логарифмической шкалы к арифметической (рис. 18, д ), мы получим картину, весьма напоминающую динамику реальных распределений (см. рис. 17). Следовательно, наше исходное предположение о том, что вместе с увеличением средних размеров особей растет и адаптируемость вида, не противоречит фактам.
Естественно, модель, которую мы здесь исследовали, весьма примитивна. Видовая дифференциация совсем не обязана всякий раз сочетаться с изменением размеров тела на какой‑то фиксированный процент. Не вызывает сомнений, что часть видообразований происходит при изменении других признаков особей, и в этом случае виды‑потомки остаются в классовых интервалах своих предков. Однако для правомочности рассмотренной модели достаточно, чтобы лишь некоторая доля всех актов видообразования сопровождалась изменением размеров тела.
Конечно, еще легче сдвиг видовых распределений вправо объяснить с позиций ортогенеза (точнее, автогенеза), т. е. действием особых сил, детерминистических законов и стремлений, заложенных в самой природе организмов. В гл. 1 мы пытались вскрыть философско‑психологические корни ортогенетических теорий. Приверженцы этих подкупающе простых идей даже вызывают симпатию своей верой в чудесные и таинственные силы природы. Однако ортогенез начисто лишен конструктивности и, по существу, несмотря на свою наукообразную форму, так же далек от объяснения эволюции, как астрология – от предсказания политических перемен.
Итак, можно сформулировать главный вопрос нашего повествования: что же лежит в основе долговременных макроэволюционных тенденций – ортоселекция или рост адаптируемости? Для ответа попробуем перейти к генетической подоплеке этих явлений.
Гены и мощность
Попробуем ответить на вопрос, как изменится величина количественного признака при увеличении числа генов, управляющих его развитием. Всякая морфологическая структура состоит из клеток и внеклеточного вещества – продукта жизнедеятельности тех же клеток. Поэтому размеры структур, органов, да и всего организма определяются числом, объемом и экспортной активностью их клеток.
Наличие гена мы определяем по его экспрессии. Если ген не активен ни в одной клетке организма, ни на одной стадии его развития, то мы можем считать, что его просто нет. Идеи о латентном (скрытом) существовании на протяжении миллионов лет вполне исправных генов ни на чем не основаны. Как селективно‑нейтральные, латентные гены должны довольно быстро терять свое информационное содержание, поскольку не проверяются отбором. В чем же выражается генная активность? Прежде всего в транскрипции РНК, хотя это лишь необходимое, но далеко не достаточное условие для генной экспрессии. Обычно РНК должна «созреть» и транслироваться в полипептид, способный узнавать другие молекулы и, исполняя свою специфичную функцию, содействовать успеху организма в его борьбе за жизнь.
Мы уже отмечали, что живое вещество можно рассматривать как катализатор, «пытающийся» ускорить собственное воспроизведение. При этом совокупное действие всех генов генома направлено на увеличение биомассы вида. Однако как быть с отдельными индивидами? Не станем же мы утверждать, что активность всех генов направлена на увеличение массы особи? Ответ здесь, казалось бы, должен быть отрицательным, но не станем спешить.
Обычно особи многоклеточных организмов диплоидны, т. е. каждая хромосома их соматических клеток представлена двумя вариантами (гомологами), хотя для многих видов растений (да и некоторых животных) можно искусственным путем получить ряды форм с измененным числом гомологов. У гаплоидов это число равно одному, у триплоидов – трем, тетраплоидов – четырем и т. д. Оказалось, что триплоиды и тетраплоиды, как правило, крупнее (мощнее) диплоидов, а последние мощнее гаплоидов. Этот эффект обусловлен прежде всего соответствующим увеличением объема клеток всех типов. Таким образом, суммарный эффект генов направлен на прирост вещества цитоплазмы клеток.
Еще в 1908 г. Р. Гертвиг установил «закон» ядерно‑плазменного отношения, гласящий, что чем больше объем ядра, тем больше объем цитоплазмы. Но объем клеточного ядра в разных тканях одного организма определяется степенью деконденсации хроматина, которая прямо связана с активностью генов. Таким образом, этот «закон» также указывает на то, что суммарный эффект работы всех генов направлен на увеличение клеточной массы.
Большинство мутаций с качественным эффектом на фенотип ведет к уменьшению размеров какой‑либо морфологической структуры или всей массы тела особи. Примерно так же влияют и микроделеции, связанные с утратой гена. Увеличение размеров структур в результате серьезного повреждения гена наблюдается гораздо реже. У многих генетически изученных объектов известен ряд локусов, потеря или серьезное повреждение которых ведет к ступенчатой редукции одной и той же структуры. Например, у гороха известна большая серия подобных генов, определяющих массу семян.
В качестве меры развития структуры следовало бы выбрать ее вклад в реализацию основных функций организма. Однако непонятно, как этот вклад измерить; гораздо проще в качестве искомой меры взять относительную величину структуры, т. е. отношение ее размера к размеру всего тела, олицетворяющего собой мощность всех функций организма.
Как мы убедились выше (см. гл.2), сложность структуры коррелирует с ее мощностью. Многочисленные складки, карманы, выпячивания обеспечивают увеличение площади функционально активных поверхностей. Естественно, каждая складка появляется в определенном месте и в определенное время и является следствием «срабатывания» конкретного гена (или группы генов). Разнообразие клеточных элементов тоже коррелирует с генетической сложностью программы развития структуры. В итоге, возникает простая логическая цепь: чем больше генов контролирует развитие структуры, тем выше ее морфологическая сложность, чем выше эта сложность, тем больше ее относительные размеры и мощность.
Все органы обладают рациональным строением, т. е. развивают максимальную мощность при минимальных энергетических затратах. Этот принцип симморфоза объясняет многократное возникновение в разных эволюционных стволах практически идентичных (в морфологическом отношении) структур, построенных как бы по одному плану. Поразительно, что тот же «план» довольно часто можно обнаружить и в конструкциях машин, созданных инженерной мыслью людей. Единственное разумное объяснение такой двойной конвергенции заключается в ограниченности спектра решений, обеспечивающих рабочую структуру максимальной мощностью при минимуме энергетических затрат.
Рассмотрим такую техническую задачу, как идентификация объекта на расстоянии. Важность ее для организмов очевидна. Внешний предмет может быть и хищником, и жертвой, и особью противоположного пола, и совершенно нейтральным объектом. Если на него падает свет, то есть одно универсальное решение – зарегистрировать особенности отражения лучей от отдельных частей объекта. Очевидно, что для этого необходимо иметь светочувствительную структуру и устройство, направляющее на него отраженный свет. Всем этим снабжены многие оптические приборы, например фотокамера.
Создавая такие приборы, человек руководствуется законами геометрической оптики и свойствами доступных ему материалов. В качестве светочувствительной структуры он использует фотоэмульсию, и фокусирует на нее лучи с помощью преломляющих свет прозрачных тел – прежде всего линз. Для получения четкого изображения объекта на фотоэмульсии мы должны уметь изменять или величину зазора между нею и линзой, или фокусное расстояние последней. Кроме того, мощность светового потока также нужно плавно регулировать, для чего оптические приборы снабжаются диафрагмой.
Животные решают задачу идентификации удаленного предмета точно так же. Глаз с высокой разрешающей способностью обладает слоем светочувствительных клеток – сетчаткой – и всеми устройствами, которыми снабжен хороший оптический прибор: диафрагмой в виде радужины, системой линз (роговица, хрусталик, стекловидное тело), мышцами, способными изменять или фокусное расстояние линз, или их отстояние от сетчатки. Мощность органа зрения, таким образом, прямо зависит от светосилы системы его «линз», количества светочувствительных клеток сетчатки, эффективности устройств фокусирования лучей и регуляции интенсивности светового потока, а также от ряда других технических моментов.
«Мощные», т. е. зоркие глаза устроены удивительно сходно у таких разных групп животных, как головоногие моллюски и высшие позвоночные. Орган зрения эти группы приобрели совершенно независимо, поскольку их эволюционные пути разошлись не позднее раннего кембрия от общего предка, который вместо глаз имел в лучшем случае лишь рассеянные по телу светочувствительные клетки. Соображения некоторых эволюционистов о возникновении глаз под действием неких внутренних факторов, толкающих орган зрения по пути прогресса, в настоящее время выглядят по меньшей мере анахронизмом.
Дата добавления: 2016-01-26; просмотров: 841;