Глава IV. Характер, особенности и причуды географического распространения
Мы рассмотрели в предыдущей главе, как (в географическом и экологическом отношении) распространены мохообразные. Но в том, что касается географии, рассматривался весь отдел в делом. Сейчас же попробуем описать характер географического распространения видов бриофитов.
Споры мохообразных мелки: минимальные их размеры составляют около 6, максимальные – около 200 микрон. Надо, впрочем, заметить, что споры диаметром свыше 100 микрон известны у представителей всего лишь двух родов. Наиболее частые размеры спор в пределах 10–20 микрон. Естественно, такие споры необычайно легки.
Крайне мелкие размеры – не более нескольких миллиметров – имеют и большинство органов вегетативного размножения этой группы растений – выводковые почки, выводковые тела и тому подобные. Некоторые из них всего лишь в несколько десятков микрон, то есть они немного крупнее спор. Понятно, что вес и этих органов тоже очень невелик.
Значит, и споры, и большинство органов вегетативного размножения бриофитов могут далеко разноситься ветром, воздушными течениями.
Добавим к этому, что споры многих мохообразных очень устойчивы к неблагоприятным внешним условиям. В опытах ряда исследователей споры печеночников и мхов прорастали после пребывания в гербарии от одного года до 5–13 лет. 13 лет выдержали хранение в гербарии и не потеряли жизнеспособности споры широко распространенного мха Funaria hygrometrica и 16 лет Ceratodon purpureus. 12 лет выдержали споры Dicranum polysetum, 11 лет – споры Trichodon cylindricum. Известен, кроме того, случай успешного проращивания спор печеночника Riella после 25‑летнего пребывания их в гербарии. Есть также данные об успешном проращивании спор некоторых мхов, пролежавших в гербарии 50 лет! Впрочем, есть и прямо противоположные факты. Например, споры некоторых печеночников теряют жизненность меньше чем через час после высыпания из коробочки.
Феноменальной устойчивостью обладают споры и к низким температурам. Подсохшие естественным путем споры некоторых листостебельных мхов выдерживали температуры жидкого азота и жидкого гелия, не теряя способности к прорастанию. А это ведь ни много ни мало от −190 до −270°C! Даже не очень понятно, зачем нужна такая чудовищная морозоустойчивость: ведь в природе споры с такими низкими температурами не встречаются.
Органы вегетативного размножения такой устойчивостью, понятно, не обладают. Однако и у них она тоже велика. Выводковые тела многих видов мхов в состоянии сохранять жизнеспособность и после нескольких месяцев пребывания в гербарии и переносить температуры порядка −30, −40°C.
Количество спор, продуцируемых мохообразными, невероятно велико. Только одна коробочка, например мха Eurhynchium confertum, производит от 280 до 700 тысяч спор! А таких коробочек на одном растении по крайней мере несколько. И это еще не рекорд и не исключение. Скорее, ближе к «золотой середине». Этот вид значительно уступает, например, видам рода Buxbaumia, производящим 5 500 000 спор, и видам рода Dawsonia, которые дают около 80 миллионов спор на одну коробочку. Вот это действительно рекорд! Исключением же являются представители немногочисленного семейства Archidiaceae, в коробочках у них образуется крайне ограниченное количество спор, не превышающее 30 (чаще 8–28).
Итогом знакомства с этими и подобными фактами явилось представление о том, что из‑за дальнего и беспорядочного разноса зачатков мохообразных их распространение довольно хаотично и не подчиняется (в отличие от папоротникообразных и семенных растений) четким географическим закономерностям.
Но мере того, как накапливался материал по географическому распространению видов мохообразных, становилось, однако, все более ясным, что представление это неверно. Одним из первых бриологов, показавших невозможность объяснения ареалов мхов беспорядочным разносом спор ветром, был русский бриолог А. А. Сапегин. В работе, посвященной изучению флоры мхов Крыма (она опубликована в 1910 году), он показал, что, если бы ареалы мхов формировались разносом спор ветром, флора мхов Крыма была бы другой.
Как это нередко случается в истории науки, работа А. А. Сапегина прошла не то чтобы вовсе незамеченной, но должного внимания коллег – как зарубежных, так и соотечественников, не привлекла.
Рецидивы старых представлений о хаотичном, распространении мохообразных порой и сейчас дают о себе знать, но господствующей все же является другая точка зрения. А именно – географическое распространение бриофитов так же строго закономерно, как и распространение всех остальных наземных высших растений.
Но закономерно не значит одинаково.
Хотя на распространение мохообразных действуют те же факторы, что и на остальные наземные растения, – в первую очередь климат, геологическая история страны и другие, однако проявляется это по‑разному, что связано с особенностями самих растений.
Целый ряд особенностей мохообразных – каких, мы расскажем ниже, – в совокупности приводит к очень широкому распространению большого числа представителей этого отдела. Уточним: речь идет уже не об отделе в целом, а о видах, входящих в него. Именно виды мохообразных имеют широкое распространение.
Вот несколько примеров.
Хорошо всем известный мох кукушкин лен (Polytrichum commune) распространен в. Европе, Азии и Северной Америке – от южных районов субтропиков на юге до Гренландии и Шпицбергена на севере, а также в Южной Америке (Перу и Бразилия), в Африке, на о‑ве Мадагаскар, в Австралии и на Новой Зеландии.
Orthotrichum speciosum имеет такое распространение: практически вся Европа; бо́льшая часть лежащих к северу от субтропиков территорий Азии; Северная Америка, примерно от 40° северной широты на юге до самых крайних точек на севере; Гренландия; северная часть Африки.
Распространение Plagiothecium denticulatum: едва ли не все лежащие к северу от субтропиков территории Европы, Азии и Северной Америки (включая горы субтропиков и отчасти тропиков), на севере – с широким заходом в Арктику, на Азорские и Канарские острова; Мадагаскар; Южная Америка (главным образом Перу); Тасмания (под сомнением), восточная часть Австралии.
Приведенные виды не рекордсмены. Есть такие, к которым почти без оговорок можно применить эпитет космополит. Эти виды можно встретить едва ли не в каждом отделе. Распространение их все же отличается от распространения основной массы видов соответствующей группы. А вот список видов мохообразных, имеющих такое же или почти такое же распространение, как и те, что названы выше, можно очень долго продолжать.
В средних и высоких (субарктических и арктических) широтах подавляющее большинство видов мхов и печеночников, свыше 90% (!), распространено циркумполярно, то есть встречается и в Европе, и в Азии, и в Северной Америке. Южнее, в тропиках и субтропиках, доля широко распространенных пантропических и пансубтропических видов, значительно ниже. Но и там мохообразных с такими ареалами немало.
К этому надо добавить, что значительное число видов встречается одновременно и в Северном, и Южном полушарии. И не только в тропиках – к югу и северу от экватора (таких видов много и среди остальных высших растений), но и в умеренных широтах обоих полушарий, с разрывами в ареалах, приходящимися на тропическую и часто субтропическую зоны.
В итоге мхи и печеночники территорий, расположенных в средних и высоких широтах, можно не без успеха определять по руководствам, составленным для стран, хотя и лежащих примерно на тех же широтах, но очень удаленных. Например, при определении мохообразных Сибири вы можете пользоваться определителями мохообразных для Северной и Средней Европы или Северной Америки. Ни в одной другой группе наземных растений такого положения нет. Конечно, это удобно. Но, может быть, это делает работу бриолога скучной? Ведь всюду сплошное однообразие. Одни и те же виды и не одиночно, в порядке исключения, а во множестве, «пачками», десятками и сотнями произрастают в лесах Канады и Сибири, на Аляске и в Северной Европе, в Восточной Азии и в Северной Америке.
Скучно? Нет. Сходство бриофлор, даже весьма удаленных друг от друга районов, велико и не может не удивлять. Можно даже сказать, оно потрясает, оно абсолютно несопоставимо со сходством во флорах остальных высших растений тех же территорий. Но именно сходство, но... не тождество!
К тому же это сходство по‑настоящему велико, если рассматривать бриофлоры территорий, находящихся примерно на тех же или близких широтах и с одинаковой степенью удаления от берега моря. Если же пойти на север или на юг или попытаться сравнить бриофлоры приморских и континентальных регионов, картина резко изменится. Степень сходства и здесь будет гораздо выше, чем во флорах остальных высших растений этих же территорий, но ясно проступят и различия. Бриофлоры, например, сибирской тайги и хвойно‑широколиственных лесов южной части советского Дальнего Востока различаются весьма существенно. И не только они. Чем дальше друг от друга отстоят по широте и степени континентальности взятые для сравнения территории, тем все бо́льшие различия в их бриофлорах мы увидим.
Так что об однообразии говорить не приходиться. О скуке – тоже.
Со времен классической работы немецкого бриолога Т. Герцога «География мхов», вышедшей в 1926 году и заложившей основы географии мохообразных, считается, что ареалы видов мохообразных по размерам сопоставимы с ареалами секций и даже родов покрытосеменных и примерно равны им. Так отличается география мохообразных от географии покрытосеменных и вообще всех высших наземных растений.
Конечно, это только схема. Встречаются виды мохообразных, ареалы которых значительно меньше ареалов родов покрытосеменных, и роды мохообразных, ареалы которых меньше ареалов видов покрытосеменных. Но это исключения.
Но не только обширность ареалов видов характерна для мохообразных. Им присуща еще, как говорят бриологи, высокая степень дизъюнктивности. Что это такое?
Во всех группах растений есть виды, имеющие не сплошное, а разорванное распространение, ареалы которых состоят из двух или нескольких, не связанных друг с другом «кусков» (участков), разделенных нередко сотнями или тысячами километров суши или моря. Мохообразные характеризуются наиболее высоким (по сравнению с остальными высшими растениями) процентом таких дизъюнктивных видов.
Разрывы ареалов могут быть очень различными и по форме, и по размерам, и, естественно, по происхождению.
Часто они приходятся на сушу.
Скажем, вид распространен в Арктике, а затем после большого перерыва появляется в горах средних и даже южных широт – на Алтае, в Саянах, на Карпатах, в Альпах, на Кавказе, в горах Средней Азии. Такое распространение свойственно большой группе арктоальпийских (их еще называют арктогорными) растений, в число которых входит и много мохообразных (только мхов и печеночников, антоцеротовые в этой группе отсутствуют). Примерами таких арктоальпийских видов являются нередкие в высокогорьях Сибири Racomitrium lanuginosum, Orthothecium chryseum, Pogonatum capillare.
Другой пример разрыва. Вид распространен в Европе, продвигаясь на восток до западных предгорий Урала, затем следует перерыв, потом он появляется в некоторых районах Южной Сибири, опять большой перерыв в распространении, и, наконец, вид появляется еще раз – теперь уже на юге Дальнего Востока. Таков характер распространения многих неморальных видов, происхождение, экология и география которых в основном связаны с широколиственными и хвойно‑широколиственными лесами. Часто эти неморальные виды встречаются только в Европе и на юге Дальнего Востока, полностью отсутствуя в Сибири. Неморальные покрытосеменные растения нередко бывают представлены разными видами в Европе и на юге Дальнего Востока. Так, в Европе встречается один вид ландыша (Convallaria majalis), в юго‑восточной части Сибири и на юге Дальнего Востока – другой (Convallaria keiskei). В Европе встречается один вид печеночницы[5](Hepatica triloba), на юге Дальнего Востока (в Сибири она полностью отсутствует) – другой (Hepatica asiatica).
У мохообразных с этим типом распространения во всех изолированных участках всегда встречается один и тот же вид. Это можно проиллюстрировать рядом примеров. Так, и в Европе, и на юге Сибири, и на юге Дальнего Востока произрастают Homalia trichomanoides, Leskeella nervosa, Mnium cuspidatum.
Еще один тип разорванного ареала. Вид встречается в Восточной Азии, включая юг советского Дальнего Востока, и потом, после очень большого перерыва, вдруг... появляется на Кавказе. Именно такое распространение имеет, например, один из представителей семейства Polytrichaceae – Pogonatum inflexum.
Нередко разрывы ареалов приходятся на моря и океаны. Это относится к очень многим видам из всех трех классов бриофитов, встречающихся в Приморье на Сахалине и на Курилах, а также на островах Японии. Таких видов, во всяком случае, несколько сот.
Но есть и более интересные формы разорванных ареалов, тоже приходящихся на воду. Так, значительная группа мхов и печеночников, распространенных в Восточной Азии (нередко включая также южную часть советского Дальнего Востока), после обширного перерыва появляются на западе Северной Америки. Иногда такое распространение называют амфипацифическим.
Очень часто разрывы ареалов имеют, так сказать, комбинированный характер: часть их приходится на сушу, часть на воду. Таких разрывов большинство. Один из примеров подобного разрыва – большая группа восточноазиатско‑восточносевероамериканских видов. Они встречаются в Восточной Азии (очень многие – в пределах советской части Дальнего Востока) и в восточной (приатлантической) части Северной Америки, в восточных и юго‑восточных штатах США. В западной половине Северной Америки, более близко расположенной к Восточной Азии, все эти виды отсутствуют. Цветковые растения с таким распространением на каждом материке представлены особыми видами. Общих видов почти нет. Мохообразные иногда тоже имеют разные (хотя и близкие) виды – на разных материках, но в большинстве случаев одни и те же, общие для Восточной Азии и восточной половины Северной Америки: Buxbaumia minakatae, Anomodon thraustus, Schwetschkeopsis fabronia, Aulacomnium heterostichum.
* * *
Вариантов дизъюнкций (разобщений) ареалов мохообразных множество. Перечислять их все мы не собираемся. Названы лишь некоторые примеры. Часть их проиллюстрирована картами ареалов (рис. 11–13), на которых можно увидеть и примеры других типов дизъюнкций, не упомянутых в тексте.
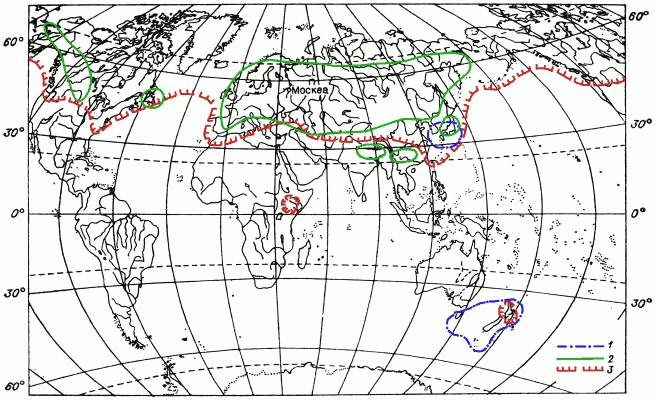
Рис. 11. Картосхема I распространения мохообразных.
1 – ареал печеночника Metzgeria furcata (пример дизъюнктивного ареала с дизъюнкцией в тропиках); 2 – ареал листостебельного мха Entodon concinnus (пример ареала широко распространенного в Северном полушарии вида); 3 – ареал листостебельного мха Hylocomium splendens (пример ареала широко распространенного в Северном полушарии вида, встречающегося в Южном полушарии).
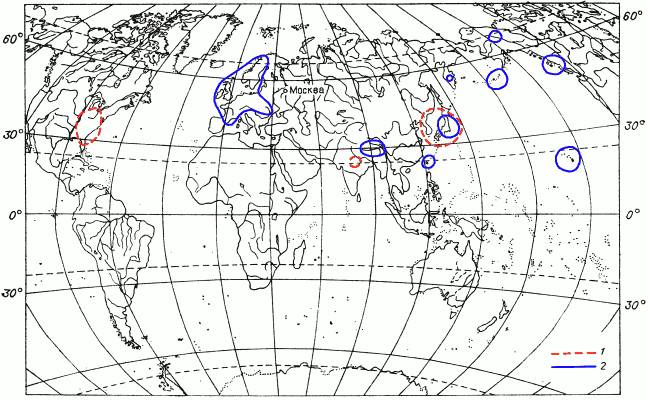
Рис. 12. Картосхема II распространения мохообразных.
1 – ареал листостебельного мха Schwetschkeopsis fabronia (пример ареала восточноазиатско‑восточносевероамериканского вида); 2 – ареал печеночника Anastrepta orcadensis (пример многократно разорванного ареала).

Рис. 13. Картосхема III распространения мохообразных.
1 – ареал листостебельного мха Pleuroziopsis ruthenica (пример ареала восточно‑азиатско‑западно‑североамериканского вида); 2 – ареал листостебельного мха Hondaella. Монотипный род (пример ареала рода, имеющего ограниченное распространение); 3 – ареал печеночника Carrpos sphaerocarpos (пример ареала вида, распространение которого ограничено Южным полушарием).
Возникает вопрос, каким образом могли возникнуть подобные ареалы. Ведь исходным для любого ареала любого вида всегда и везде является монолитность, то есть ареал, состоящий только из одного куска. Все равно, большого или маленького. Откуда же берется несколько кусков?
В чисто теоретическом плане можно предположить, по меньшей мере, три способа, три пути создания дизъюнкций. 1. Виды в каждом изолированном участке возникают всякий раз заново, независимо друг от друга. 2. Изолированность создается дальним и сверхдальним разносом зачатков мохообразных (главным образом спор) с помощью воздушных течений. 3. Раздробленный на серию изолированных участков ареал представляет собою остаток некогда единого, сплошного (монолитного) ареала.
Рассмотрим каждый из этих возможных путей.
Первый путь. Многократное возникновение вида. Исходя из того, что мы сегодня знаем об эволюции и механизме видообразования, многократное возникновение вида можно представить лишь с очень большим трудом. И уж вовсе маловероятным кажется допущение о сколько‑нибудь широких масштабах этого явления. Вообще этот путь – скорее логическое допущение, чем основанный на реалиях вывод. Короче говоря, этот путь создания разорванных ареалов мохообразных мы должны сразу же и решительно отбросить. Неприемлем он и в отношении других растений.
Второй путь. Дальний и сверхдальний разнос зачатков. Здесь надо рассмотреть по крайней мере два варианта – вид растет на суше, в глубине континента (первый вариант) и вид растет у берега моря (второй вариант). В обоих случаях разнос будет происходить преимущественно в направлении господствующих ветров (господствующих в сезон созревания спор или других зачатков). В отдельные моменты зачатки могут быть подняты на большую высоту, отнесены далеко и приземлиться на каком‑либо участке, не контактирующем с границей ареала. На военном языке это называется десантом. Оставим пока в стороне вопрос о возможной приживаемости зачатков вдалеке от основного ареала. Важно отметить, что такой заброс зачатков на удаленный от местопроизрастания вида участок возможен.
Но если рассматривать разнос зачатков не как однократный акт, а как процесс, идущий непрерывно в течение всей жизни вида, а она исчисляется миллионами, десятками миллионов, а подчас, возможно, и сотнями миллионов лет, то станет совершенно очевидным, что такой разнос не может привести к формированию изолированных участков ареалов. Разнос зачатков, рассматриваемый во времени, в целом обратно пропорционален расстоянию. Чем дальше от растения, произведшего зачатки, находится участок, тем меньше на него попадет зачатков. И наоборот. Значит, наращивание, прирост ареалов должны происходить постепенно, шаг за шагом, без образования оторванных от основного участка фрагментов. Другими словами – сплошным фронтом.
Такой прирост будет происходить до естественной преграды. Преградой может служить, например, море. Здесь прирост ареала прекращается. А если там, «за синим морем», лежащим в направлении господствующих ветров, лежит остров или материк с условиями, более или менее соответствующими данному виду? Могут ли в этом случае диаспоры попасть туда, прижиться на новом месте и, таким образом, привести к возникновению разорванного ареала?
Чтобы ответить на этот вопрос, обратимся к результатам чрезвычайно интересных опытов голландского бриолога ван Зантена [Van Zanten, 1978], решившего проверить, существует ли связь между степенью устойчивости спор мхов к неблагоприятным воздействиям и характером географического распространения видов. Ученый исследовал споры ряда видов мхов, произрастающих в Южном полушарии, в отношении устойчивости к низким температурам и высушиванию. Именно устойчивость к названным факторам и определяет жизненность спор в условиях дальней и длительной транспортировки воздушными течениями. Споры одних и тех же видов подвергались суровым испытаниям в различных режимах: высушивались, часть их после этого выдерживалась в условиях низкой температуры (−30°C) в течение различного времени, часть увлажнялась. После этого споры проращивались. По проценту спор, не потерявших способности прорастать, можно было судить об устойчивости тех или иных видов.
Результаты этих исследовании ван Зантен сопоставил с данными по географическому распространению видов, споры которых исследовались. Устойчивость их была, если можно так выразиться, «наложена» на карту ареалов.
И вот что оказалось. Опыты показали, что существует ясная и несомненная связь между степенью устойчивости спор и размером ареалов видов. Чем более устойчивы споры у того или иного вида, тем обширнее у него ареал. Наиболее устойчивые споры имеют виды, распространенные в Новой Зеландии, Австралии (обычно вместе с Тасманией), в Южной Америке, а также и на антарктических островах, то есть имеют максимально широкое для Южного полушария распространение. И наоборот, виды с менее устойчивыми спорами имеют более ограниченные ареалы, отсутствуя на каких‑либо (из тех, что были названы) территориях. Наименее устойчивыми к низким температурам и высушиванию оказались споры у эндемов Новой Зеландии[6].
Что это? Случайность? Вряд ли. Вывод может быть только один – на разделенных морем участках суши разорванность ареалов создается посредством дальнего, очень далекого (на несколько тысяч километров) разноса спор. Именно такой вывод и делает ван Зантен.
Таким образом, видим, что дальний разнос спор может формировать у мохообразных разорванные ареалы. Однако таких случаев мало. А непременным условием для создания такого ареала должно быть наличие морей или океанов между изолированными участками и размножение спорами, так как только, они способны сохранять жизнеспособность после всех невзгод длительных трансокеанских путешествий. У большинства же видов размножение спорами стоит на втором месте после вегетативного. Очень многие виды, в том числе и такие, что встречаются на разных континентах, спорами не размножаются.
Значит, для объяснения происхождения большей части разорванных ареалов мохообразных этот путь – дальнего и сверхдальнего разноса спор – непригоден. Он не может объяснить причин возникновения всех разорванных ареалов, например: разрывов, приходящихся на сушу, разрывов ареалов видов, размножающихся исключительно вегетативным путем и т. д. Кроме того, не надо забывать, что спорам, волею судеб оказавшимся вдали от основного ареала, во‑первых, очень трудно попасть на участок с подходящими условиями, во‑вторых, очень трудно удержаться на них. Все‑таки чем дальше от «родного дома», тем больше всякого рода трудностей, тем меньше шансов у далеко заброшенного «десанта» захватить плацдарм и удержать его. Как правило, такие «десанты» обречены на гибель.
Остается третий путь, рассматривающий разорванные ареалы как остатки некогда единого, монолитного ареала.
Каким образом единый, цельный ареал распадается на два или несколько не связанных между собою участков, кусков, отстоящих один от другого нередко на тысячи километров? В схеме – очень просто: вид когда‑то занимал обширную площадь, но затем под влиянием неблагоприятных условий на значительной части ее вымер (во всяком случае исчез), оставшись на двух или нескольких участках, где условия почему‑либо изменились меньше. Если эти участки далеко друг от друга, то и возникает разорванный, дизъюнктивный ареал.
Обратимся к одному из наиболее понятных разрывов ареалов – арктоальпийскому. Напомним, что арктоальпийские виды обитают, с одной стороны, в Арктике, а с другой – в горах (главным образом в высокогорьях) средних и даже сравнительно южных широт, например на Кавказе или на Тянь‑Шане. В эпохи оледенений арктические ландшафты и флора арктического типа продвигались далеко на юг.
В то же время нижняя граница безлесных высокогорных ландшафтов опускалась во многих случаях до подножий горных хребтов. Граница между продвинувшейся далеко к югу флорой арктического типа и резко снизившейся альпийской флорой стала очень узкой, а во многих случаях вообще исчезла. Произошло смыкание двух различных флор; И между ними начался интенсивный обмен видами. В бриофлорах, где альпийских видов было мало, происходила, вероятно, преимущественно экспансия арктических видов в пределы горных территорий. Виды, которые мы сейчас называем арктоальпийскими, были распространены от Арктики до гор средних и южных шпрот сплошной полосой. Ареалы их были едины и монолитны.
По вот наступило потепление. Холодолюбивым видам (будущим арктоальпийцам) стало неуютно. Обособилась и постепенно расширилась граница между северной и южной частями ареалов. Южная граница видов арктической флоры отодвинулась на север, а нижняя граница этих же видов, но растущих в горах, стала подниматься вверх. Обширные равнинные территории, нижние и средние части гор для этих видов стали непригодными и заселились несколько более теплолюбивыми, главным образом таежными видами. В итоге первоначально единый сплошной ареал раскололся на арктическую часть и серию изолированных от нее и друг от друга высокогорных участков, расположенных в горах средних и южных широт.
Описание этого процесса мы несколько схематизировали. В частности, пренебрегли тем обстоятельством, что эпох оледенения и межледниковья было несколько. Значит, весь процесс дробления и последующего слияния ареалов неоднократно повторялся.
Это – один из примеров, характеризующих преобразование ареала из единого во множественный. Вариантов дизъюнктивных ареалов, как мы отмечали, много. Каждый требует конкретного объяснения. Общее же во всех случаях одно – изменение условий приводит к исчезновению вида с части занимаемой им территории, в то время как на другой части (или в других частях) с более благоприятными условиями он сохраняется.
Даже в эпоху максимального оледенения Арктика не представляла собой сплошь покрытого льдом пространства. Были участки – и нередко значительные по площади – свободные ото льда. На них, несмотря на суровость климата, сохранялась растительная жизнь. И сейчас в некоторых районах Арктики (широкой известностью в этом отношении пользуется, например, северная часть Аляски) сохранились виды мохообразных, росшие там еще до того, как флора Арктики стала арктической в сегодняшнем понимании. Арктические местонахождения этих видов далеко оторваны от основных участков ареалов, расположенных значительно южнее, в умеренной зоне.
Таким образом, приходим к выводу, что главный путь образования разорванных ареалов – это дробление первоначально цельного, единого ареала, связанное с изменением условий, в первую очередь климата. Другие пути формирования таких ареалов, например дальний разнос спор, имеют лишь второстепенное значение.
Отсюда – важность, огромная важность изучения разорванных ареалов, в частности, мохообразных для познания истории изменений природных условий, истории флоры и растительности того или иного региона.
Дата добавления: 2016-01-26; просмотров: 1761;