Глава III. Условия жизни
Широко бытует мнение – не миновало оно и ботаников – будто мохообразные растут главным образом во влажных или сырых местах, то есть что эти растения по преимуществу северо‑болотно‑горные и лесные. Да, бриофиты действительно в изобилии растут и на болотах, и в лесах, и на севере, и в горах. Но не только там.
Мохообразные почти вездесущи. Они встречаются на всей территории земного шара. Географический диапазон их поистине необозрим. От экватора до высоких широт Арктики и Антарктики, от уровня моря до высочайших горных вершин. Нет территории, где мохообразные вовсе отсутствовали бы. В одних случаях они играют большую роль в сложении растительного покрова, в других – незначительную, но всегда и везде мохообразные – неотъемлемый и непременный компонент растительного мира региона. Никакое ботаническое изучение территории, все равно лесной или пустынной, болотной или тундровой, не будет поэтому полным без учета бриофлоры, представители которой всегда (подчеркиваем, всегда) принимают участие в формировании растительного покрова.
Чрезвычайно широк и экологический спектр мохообразных. Если рассматривать весь отдел в целом, то кажется, что для него нет непреодолимых барьеров, неприемлемых, неподходящих условий.
Какой бы экологический фактор мы ни взяли, увидим, что мохообразные встречаются в пределах всей полосы изменений этого фактора, вплоть до его крайних значений, при которых еще возможна жизнь высших растений, а в ряде случаев вообще растительная жизнь.
Если этот фактор – влажность, то на одном полюсе его мы встретим мохообразные, живущие в воде, а на другом – на почти полностью лишенных воды местообитаниях.
Бриофиты, живущие в воде, распадаются на две большие группы: свободно плавающие на поверхности и полностью погруженные. Первых сравнительно немного. Это главным образом печеночники. Из них наиболее распространены и известны Ricciocarpus natans и Riccia fluitans. Вторых гораздо больше – многие десятки видов. Эти виды закреплены в донном грунте и нередко идут на значительную, подчас огромную глубину – почти до 60 метров. Именно на такой глубине была зафиксирована одна из форм довольно широко распространенного и, между прочим, наземного вида Thamnobryum alopecurum. Обнаружен был этот мох в воде Женевского озера в ту пору, когда вода в нем еще славилась своей чистотой и прозрачностью. Как обстоит дело сейчас, мы не знаем.
На глубинах 32 и 33–36 метров в озерах Восточной Антарктиды участниками советских экспедиций М. И. Симоновым и В. С. Короткевич были обнаружены мхи. Изучение мхов Л. И. Савич‑Любицкой и З. Н. Смирновой показало, что это новые для науки виды из родов Plagiothecium и Bryum.
Но глубоководных видов не так уж много. Более часто пределом является глубина 10–12 метров. На таких глубинах отмечен, например, Fontinalis nitida на Селенгинском мелководье на Байкале.
Глубины же от 20–30 сантиметров до 3–5 метров обычны. Здесь произрастает большое количество видов, главным образом листостебельных мхов из семейств Amblystegiaceae и Fontinaceae. Они практически повсеместны.
Говоря о водных мхах и вообще водных мохообразных, заметим, что почти все они растут лишь в пресной воде. В морской воде бриофиты почти полностью отсутствуют. Одним из немногих исключений является Fontinalis antipyretica, встречающийся в Балтийском море, вода которого, как известно, отличается большей опресненностью. В пресной воде указанный вид распространен чрезвычайно широко. Но в то же время мхи (именно мхи) сравнительно широко распространены на прибрежных скалах, в непосредственной близости от воды, на участках, регулярно орошаемых брызгами морских волн. Таких видов во всяком случае несколько десятков. Не вынося полного погружения в морскую воду (это характерно для большинства высших растений), многие виды бриофитов спокойно переносят обильное и постоянное увлажнение ею.
Полная и постоянная погруженность в воду – это одно крайнее значение фактора влажности, один полюс. Другой – почти лишенные воды местообитания – пустыня. Да, мохообразные растут и в пустынях! Не так уж их там много, но есть. Один из наиболее обычных и распространенных пустынных мхов – Tortula desertorum. Он часто встречается в песчаных пустынях, покрывая местами территорию на 40–70 и даже на 90%! О том, насколько этот вид массовый и какое значение он имеет в образовании растительности Средней Азии, можно судить по такому факту. Ассоциации с господством Tortula desertorum так широко распространены, что даже нашли отражение на геоботанических картах, и не только на крупномасштабных. Так, в «Атласе Узбекской ССР», например, на картах растительности показаны ценозы с господством пустынной тортули.
Кроме собственно пустынь мохообразные, главным образом листостебельные мхи из семейств Pottiaceae, Trichostomaceae и Grimmiaceae, растут на скалах в пустынных областях. Влаги здесь так же мало.
Ну и, конечно, мохообразные встречаются (и часто в изобилии) на всех возможных переходных территориях от воды до безводных пустынь и сухих пустынных скал.
Обратимся теперь к такому важному экологическому фактору, как свет. И здесь – на одном полюсе максимальная и постоянная освещенность (в тех же пустынях и на пустынных скалах), а на другом – тоже очень большая и постоянная затененность, например в тропическом лесу или в пещерах. В тропических лесах бриофиты растут в условиях освещения примерно в 150–200 раз более слабого, чем полное освещение тех же широт.
В условиях крайне слабого освещения, чуть ли не в полной темноте, растут листостебельные мхи в пещерах. В 1977 году дальневосточный бриолог В. Я. Черданцева и автор настоящей работы, исследуя Макрушинскую пещеру, находящуюся в Южном Приморье, обнаружили там несколько видов листостебельных мхов. Мхи Timmia megapolitana и Mnium stellare встречались на расстоянии до 20 метров от входа, а максимальное удаление от входа Amblystegiella jungermannioides составило 30 метров. Без фонариков передвигаться в пещере было невозможно. Но нельзя сказать, что названные мхи выглядели угнетенно. Нет. Обычные, нормально развитые экземпляры. Заметим, что какие‑то специально пещерные мхи среди них отсутствовали. Впрочем, их и на всем земном шаре вряд ли наберется больше 3–5 видов. Все перечисленные виды преимущественно встречаются вне пещер. И в нашем случае Timmia megapolitana росла не только в пещере, но и возле входа в нее, с наружной стороны.
В условиях постоянного недостаточного освещения у некоторых видов мхов выработались чрезвычайно своеобразные приспособления для улавливания самых скудных количеств лучистой энергии, которые достигают дна пещер. Широко известным в этом отношении, можно сказать, классическим примером является Schistostega pennata. Она имеет долгоживущую протонему, пластинка которой всегда ориентирована перпендикулярно направлению лучей света. Состоит пластинка из линзовидных клеток, которые преломляют лучи света и фокусируют их на сконцентрировавшиеся возле задней (по отношению к источнику света) стенки хлоропласты. Часть света при этом отражается от задней стенки клеток протонемы (рис. 10). В результате протонема светится или, точнее, поблескивает нежным золотисто‑зеленым светом. Он очень хорошо заметен в полумраке пещер, в затененных нишах среди скал или в густых хвойных лесах – словом, там, где растет Schistostega pennata.
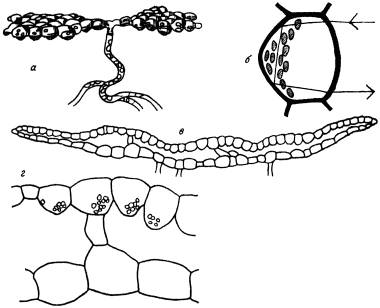
Рис. 10. Детали строения «светящихся» мохообразных.
а – пластинчатая протонема листостебельного мха Schistostega pennata; б – отдельная клетка протонемы в поперечном разрезе со схематическим изображением хода лучей, заштрихованные тельца в левой части клетки – хлорофилловые зерна; в – поперечный срез слоевища печеночника Cyathodium smaragdinum; г – фрагмент среза, сильно увеличенный. Хорошо видно большое сходство формы клеток верхнего слои печеночника с формой клеток протонемы схистостеги. Хлорофилловые зерна у печеночника, как и у схистостеги, концентрируются у задней стенки клеток.
Свечение это чрезвычайно эффектно и производит сильное впечатление.
Свечение протонемы Schistostega pennata известно народам Европы (где этот вид распространен сравнительно широко) с очень давних пор. Оно послужило источником, основой для множества легенд и мифов, рассказывающих о таинственных золотых кладах, не дающихся в руки.
Вот что писал об этом А. Кернер [1903, с. 385–386] в знаменитой книге «Жизнь растений»: «Это свечение, прекращающееся в тот же момент, когда светящийся предмет выносят из пещеры на яркое солнце, настолько поражает наблюдателя, что становится понятным, откуда произошли сказки о шаловливых гномах, о пещерных духах, показывающих жадным смертным золото и алмазы. Восхищенные кладоискатели с восторгом принимаются за работу, но гномы готовят им сильное разочарование: когда открывают мешки с нарытым второпях пещерным кладом, то вместо самоцветных камней из них вываливается грязная земля».
Схистостега неодинока в своем необыкновенном «умении» действовать подобно увеличительному стеклу, фокусируя рассеянный свет, часть которого при этом отражается и создает эффект свечения. Не так давно выяснилось, что этими способностями, основанными на очень похожем строении протонемы, обладает австралийский мох Mittenia plumula. Растет этот вид в условиях, аналогичных тем, при которых растет схистостега. Протонема его долгоживуща и тоже светится.
Известно это явление и среди печеночников. По крайней мере, два вида Cyathodium cavernarum и близкий к нему C. smaragdinum обладают способностью «светиться» вследствие преломления и отражения попадающих на поверхность слоевища лучей света. Клетки поверхностного слоя прозрачные и линзовидные, по форме очень похожие на клетки протонемы схистостеги (см. рис. 10). Встречаются эти «светящиеся» печеночники в пещерах, главным образом в тропиках, но один из них – Cyathodium smaragdinum отмечен и за пределами тропиков, на юге Японии (о‑в Кюсю).
Не является для мохообразных препятствием и меняющееся в широких пределах в течение вегетационного сезона освещение. Именно такое освещение свойственно широколиственным лесам. Весной и осенью здесь много света, летом – глубокая тень. Бриофитов в широколиственных лесах, притом самых разнообразных, уйма.
Не составляет исключения и температура. Коротко охарактеризованное выше географическое распространение мохообразных показывает, что амплитуда значений температуры, при которой растут мохообразные, так же велика, как и амплитуда других экологических факторов. Устойчивость к очень различным значениям температуры явилась одним из важнейших моментов, обеспечивших этой группе растений широкое распространение по всему земному шару.
Вот почему мы находим мохообразные и в экваториальных лесах, и в холодных арктических и антарктических пустынях, и близ линии вечных снегов на высочайших горных системах мира.
Одним из «трудных» местообитаний, мало пригодных для жизни растений, являются вода и берега термальных источников, особенно обильных и мощных в районах, где активно проявляется вулканическая деятельность. В нашей стране это главным образом Дальний Восток: Камчатка, Курильские острова, отчасти Чукотка. И здесь, на берегах горячих ключей и в них самих, при постоянном обильном увлажнении водой, имеющей температуру не менее 40–45°C, встречаются мхи и печеночники. Один из печеночников Solenostoma thermarum на Южных Курилах, по нашим наблюдениям, как раз и приурочен к таким условиям и нигде более (в пределах Южных Курил) не встречается. Обычно лишь по берегам горячих источников растет он и в Японии. Довольно хорошо чувствуют себя при постоянном подогреве сфагны. В Исландии некоторые из них растут в воде, температура которой может достигать 65°C! Примерно при такой же температуре они встречены и на Южных Курилах. При более высоких, достигающих 70–80°C, растут только водоросли. Высших растений нет. Но мохообразные, как видим, недалеко отстали от водорослей.
В начале главы уже шла речь о том, что мохообразные встречаются при тех крайних значениях экологических факторов, при которых еще возможна жизнь высших растений или даже вообще любых растений. При крайних значениях любого экологического фактора мохообразные идут дальше всех остальных высших растений. Часто рядом с бриофитами встречаются также водоросли и лишайники. Последние нередко идут дальше мохообразных. Так, лишайники составляют предел растительной жизни в наиболее холодных, а также наиболее сухих местах, а водоросли – в наиболее горячих источниках. Но на втором месте после них мы видим мохообразных, оставивших позади все остальные отделы растений. Именно мохообразные являются единственными представителями высших растений в ряде районов Арктики, Антарктики и в наиболее высоких горных системах.
Необыкновенно многообразны и потому трудно поддаются перечислению субстраты, на которых селятся бриофиты.
Ну, конечно, прежде всего, как всякие «порядочные», «нормальные» растения, мохообразные поселяются на почве. Но, вообще говоря, напочвенных видов бриофитов как раз очень немного. Большинство их растет на совсем других субстратах, подчас даже кажущихся неприемлемыми для растений. Но то, что непригодно для других высших растений, легко используется мохообразными.
Рассмотрим (конечно, коротко) некоторые из этих субстратов. Это не только интересно, но и поучительно и является хорошей иллюстрацией известного положения: природа не терпит пустоты.
О воде мы уже говорили. Понятно, «вода» в смысле «субстрат» звучит несколько странно, но в отношении водных растений, полностью погруженных в нее, можно, вероятно, говорить о воде именно как о субстрате. Кроме чисто водных (плавающих и погруженных) есть немало видов мохообразных, ведущих полуводный образ. Это многие болотные виды, растущие в постоянно переувлажненных местах, а также виды, растущие на берегах речек и ручьев и на камнях в их руслах. Колебания уровня воды приводят к тому, что такие виды оказываются попеременно то погруженными в воду, то полностью вышедшими из нее, то наполовину в воде, наполовину в воздухе. В таком же положении оказываются и многие виды, растущие в неглубоких, часто пересыхающих лужах.
Скалы представляют собой целый конгломерат различных субстратов, так как сложены различными горными породами, по‑разному ориентированы по отношению к странам света и т. д. На скалах есть сравнительно пологие участки, прикрытые более или менее мощным слоем гумуса. Есть мелкие расщелины и трещины, заполненные небольшим количеством гумусного, мелкоземного или щебнистого материала. И всюду поселяются мохообразные. Находят они себе место и на поверхности камней. И не только на горизонтальной, на которой всегда есть какое‑то количество гумуса или мелкозема, но и на крутонаклонной, вертикальной и даже нависающей, как бы опрокинутой. Никакого гумуса, мелкозема здесь, понятно, нет и быть не может. Голый камень. Но и это не препятствие для мохообразных. На совершенно обнаженной поверхности камня тоже растут мохообразные, чаще всего листостебельные мхи. Целый букет семейств имеет здесь своих представителей и нередко массовых. Наиболее широко представлены семейства Grimmiaceae, Trichostomaceae и Pottiaceae.
Растущие на поверхности камней мхи имеют в основном две формы роста – мелкие кустики из прямостоячих или приподнимающихся стеблей и ветвей и плотно прижимающиеся к субстрату коврики‑дерновинки из стелющихся по субстрату и чуть ли не вдавленных в него стеблей с отходящими от них более или менее перпендикулярно ветвями. В обоих случаях растение отыскивает и использует крохотные, подчас микроскопически мелкие углубления в камне, в которых скапливаются опять же почти микроскопические количества гумуса или хотя бы пыли. Обнаженная поверхность камней только для невооруженного глаза кажется однородной, гладкой и плотной. На самом деле ее верхний слой толщиной, может быть, всего в несколько миллиметров всегда несколько нарушен. Он пористый. Между частицами камня есть крохотные пустоты. Они заполнены воздухом, пылью, растительными остатками. Сюда‑то и внедряются ризоиды мхов. Вот почему растущие на поверхности камней бриофиты не удается вполне отделить от субстрата – или растение обрывается или к нему пристают мелкие, как песчинки, частички разрушенного камня.
На поверхности камня, где растение испытывает дефицит влаги и питательных веществ, условия для его жизни крайне неблагоприятны. Не случайно только мохообразные и лишайники могут здесь найти себе приют.
Велико разнообразие субстратов, на которых поселяются мохообразные в лесах. На почве, как уже говорилось, растет сравнительно небольшое число видов. Но зато в хвойных (в так называемых моховых) лесах напочвенный покров, образованный мхами и прежде всего такими видами, как Hylocomium splendens и Pleurozium schreberi, может создавать сплошной ковер мощностью (высотой) в 10–12 сантиметров, а нередко до 15–20. Простираются такие леса в таежной полосе, особенно в Сибири, на десятки, сотни и тысячи километров. Сплошной моховой покров бывает развит также в горных тропических и субтропических лесах.
Широко используют мохообразные в качестве субстрата для поселения гниющую древесину. Ее всегда много в любом лесу. Это пни, опавшие ветви, а главным образом лежащие на земле стволы деревьев, упавших от старости или чаще вывороченных ветром. Таких деревьев особенно много в горных районах, где слой почвы довольно тонок и корневая система имеет поверхностное расположение.
В зависимости от степени разложения древесины меняется видовой состав поселяющихся на ней мохообразных. Одни виды тяготеют к сравнительно свежей, только начинающей гнить древесине, другие, напротив, предпочитают последние стадии разложения, когда древесина становится очень рыхлой, легко рассыпается и постепенно смешивается с почвой. Третьи занимают промежуточное положение. С гниющей древесиной связаны сотни различных видов листостебельных мхов и печеночников из самых различных порядков и семейств. Объединяет их всех частичный сапрофитизм и более или менее ясно выраженная мезофитность[4]. Называются они эпиксилами.
Еще один субстрат, заселяемый мохообразными в лесу, – стволы и ветви живых деревьев и кустарников. Поселяющиеся на них мохообразные называются эпифитами.
Эпифитность широко распространена в растительном мире. Вероятно, каждый наслышан об эпифитных тропических орхидеях, об эпифитных папоротниках и различных других, произрастающих главным образом в тропиках и отчасти в субтропиках.
Эпифит использует другое растение лишь как место прикрепления, но не живет за его счет. Это не паразит. Он использует гумус, накапливающийся в трещинах коры дерева, влагу, стекающую по стволу во время дождя или выпадающую в виде росы.
Два фактора резко ограничивают распространение эпифитов – температура и влажность. Сидящий на стволе дерева в нескольких метрах от земли эпифит, как правило, не может быть прикрыт на зиму сколько‑нибудь мощным слоем снега и вынужден зимовать, так сказать, «на открытом воздухе», без снежной шубы, которая, как известно, хорошо защищает растения от холода. Это резко уменьшает шансы на произрастание эпифитов в холодных областях, например в таежной полосе. И здесь мы видим полное отсутствие эпифитов среди цветковых растений и почти полное – среди папоротников. Мохообразные же как растения значительно более выносливые сохраняются. Но здесь наблюдаются многие особенности по сравнению с тропическими и субтропическими лесами, даже с широколиственными лесами умеренной зоны.
В тайге мохообразных эпифитов мало. Мало и в том смысле, что видов небольшое число, и в том, что обильного, массового развития они не имеют. Кроме того, очень заметно снижен верхний предел, до которого эпифиты поднимаются по стволу дерева. Резко тяготеют они к основаниям и нижним частям стволов, то есть к участкам, которые на зиму закрываются снегом. Наши наблюдения показали, что большинство мохообразных эпифитов, встречающихся в тайге, зимуют под снегом.
Ряд видов, которые в более южных и соответственно более теплых краях растут в лесах в качестве эпифитов, в тайге переходят со стволов и ветвей деревьев на скалы, поверхность камней. Это позволяет им зимовать под снегом.
Эпифиты почти всю необходимую для них влагу получают из атмосферы (дождевая вода, роса, туманы), а не из субстрата, почти лишенного влаги. Поэтому водоснабжение для них по степени влажности стоит на первом месте. И хотя большинство эпифитов, во всяком случае эпифитных мохообразных, весьма скромны в потреблении влаги, все же совсем без воды и они обходиться не могут. Отсюда резкое и безусловное тяготение эпифитов к влажным районам. В тайге влажные районы важны для эпифитов вдвойне – летом они обеспечивают их водой, зимой – закрывают снегом. Снеговой покров во влажных районах более мощный, рано устанавливается и поздно разрушается.
В районах наиболее массового и обильного развития эпифитов (влажные тропические и субтропические леса) они встречаются на подавляющем большинстве стволов деревьев, густой шубой закрывая весь периметр ствола, и идут от его основания до первых ветвей, нередко более или менее далеко проникая в глубь кроны. В особенности много эпифитов на верхней стороне наклонных ветвей. Верхний предел распространения их проходит в этих случаях на высоте нескольких десятков метров от земли.
В широколиственных и смешанных лесах умеренной зоны эпифитов меньше и по числу видов, и по массе. Но и здесь в наиболее благоприятных местах (например, на Кавказе и в некоторых районах юга советского Дальнего Востока) эпифиты еще очень обильны. Они могут встречаться почти на каждом стволе дерева. В виде прерывистых полуцилиндрических лент (охватывающих примерно половину периметра ствола) эпифиты часто поднимаются высоко вверх по стволу, нередко до первых ветвей, на высоту, превышающую 8–12 метров.
Пожалуй, наиболее замечательны среди эпифитов и производят наиболее сильное впечатление свисающие формы. Они напоминают хорошо известные свисающие лишайники‑бородачи из рода Usnea, только не бывают никогда такими длинными. Длина прядей эпифитных мхов, как правило, не превышает 30–40 сантиметров, обычно меньше. Они чрезвычайно колоритны и придают лесу какую‑то мрачноватую таинственность. Появление в таком лесу сказочных персонажей воспринялось бы как нечто вполне естественное.
Встречаются свисающие эпифиты главным образом в тропиках и субтропиках. Изредка заходят они в пределы распространения широколиственных лесов умеренной зоны. В Советском Союзе свисающие эпифитные мхи можно встретить на Кавказе – на Черноморском побережье, в Ленкорани, а также на юге Дальнего Востока – в южной части Приморского края и на Южных Курилах. В Южном Приморье свисающие пряди образуют три вида: Leucodon pendulus, Haplohymenium flagelliforme и иногда Boulaya mittenii. На Курильских островах встречается последний вид.
Число эпифитных видов мохообразных на земном шаре велико и составляет несколько сот, вероятно, значительно больше 500. Эпифиты есть среди представителей всех трех классов мохообразных.
И совсем уж удивительный субстрат, на котором тоже поселяются мохообразные, – листья. Разумеется, листья вечнозеленых растений: у листопадных срок жизни листьев короток, – всего один сезон, большинство же мохообразных – многолетники.
Большинство живущих на листьях мохообразных (они называются эпифиллами) – печеночники, среди них преобладают представители семейства Lejeuneaceae. На земном шаре эпифилльных видов, вероятно, несколько сот. Только на юге Китая, по данным китайского бриолога П. Чена, насчитывается 71 вид эпифилльных печеночников.
Так как вечнозеленые растения в основном жители тропиков и субтропиков, то и эпифиллы тоже обитатели далекого юга. Так, во всяком случае, считалось до последних лет. Но в 1973 году канадские бриологи Д. Витт, М. Остафичук и И. Бродо опубликовали сенсационное сообщение о находке эпифилльных мохообразных далеко за пределами не только тропиков, но и субтропиков, на 49° северной широты. Любопытно, что эпифиллами оказались не какие‑либо экзотические виды, а довольно обычные, сравнительно широко распространенные в Канаде в качестве эпифитов. Это были три вида листостебельных мхов из рода Orthotrichum и один печеночник. Один из листостебельных мхов Orthotrichum speciosum широко распространен и в Советском Союзе. Росли все эти виды на хвое туи Thuja plicata. Вечнозеленых лиственных деревьев в тех краях нет.
После этой находки можно было предположить, что. эпифиллы, вероятно, произрастают и в других районах умеренной зоны, по крайней мере в наиболее влажных и теплых, но пока там не выявлены. Одним из таких районов является юг советского Дальнего Востока. Здесь и тепло и влажно. Автору довелось там работать в течение ряда лет. Совместно с В. Я. Черданцевой мы предприняли специальные поиски эпифиллов. Наиболее вероятным нам казалось обнаружение их на листьях таких хвойных деревьев, как пихта цельнолистная и тис остроконечный, а также на листьях вечнозеленых лиственных кустарников скиммии, падуба и некоторых других. Искали и на материковой части, и на островах – Попова, Петрова, Южном Сахалине, Итурупе, Кунашире, Шикотане. Как ни странно, наши поиски не дали результатов. Эпифиллов мы так и не нашли. Не хочется думать, что плохо искали. И в то же время как‑то не очень верится (после находки канадских бриологов), что эпифиллов действительно нет на Дальнем Востоке. Как знать, может быть, последующим, бриологам повезет больше, и они найдут эпифилльные мхи или печеночники. Острова юга советского Дальнего Востока кажутся, несмотря на неудачу наших поисков, все же довольно перспективными в этом отношении.
Еще одно удивительное местообитание. Тоже лесное. Листья многих тропических эпифитов из семейства бромелиевых (это покрытосеменные растения) смыкаются своими основаниями, образуя воронку, в которой всегда находится вода. Вы думаете, мохообразные «прошли мимо» такого удобного местообитания? Ничего подобного. В воде воронок тоже живут мохообразные: мох Philophyllum tenuifolium.
Мы привели немало примеров довольно неожиданной экологии мохообразных, проявляющейся, в частности, в субстратах, на которых эти растения поселяются.
Но, пожалуй, наиболее экстравагантным субстратом из всех, что использованы бриофитами, являются раковины живых (заметьте: живых!) пресноводных моллюсков. Это довольно крупные двустворчатые моллюски из североамериканских родов Quadrula и Lampsilis, несколько напоминающие нашу беззубку. Именно на них поселяются, живущие в воде Fissidens fontanus и Leptodictyum riparium. Прямо как актиния на раковине рака‑отшельника. Но актиния все‑таки животное, хотя и неспособное к самостоятельному передвижению, а тут растения! Зачем им надо передвигаться? Кажется, эти два вида из родов Fissidens и Leptodictyum единственные среди всех растений (высших – во всяком случае), местообитания которых способны самостоятельно передвигаться.
Кроме раковин моллюсков, этот вид фиссиденса растет еще и на другом животном субстрате, правда, неподвижном, – пресноводных губках. А еще один вид из этого же рода – Fissidens brachypus – растет уже исключительно на губках (тоже пресноводных), причем только одного вида – Metania reticulata. Обитает эта пара в тропиках Южного полушария, бассейне Амазонки. Леса, в которых живут губка и мох, ежегодно на несколько месяцев заливаются водой. И мох, и губка оказываются в нее погруженными.
И еще пример очень своеобразного субстрата и соответственно очень своеобразной экологии. Это семейство Splachnaceae. Представители его растут на субстратах животного происхождения. Это – экскременты самых различных животных, как диких, так и домашних, хищных и травоядных, погадки (отрыгнутые хищными птицами комочки непереваренной пищи). Все эти субстраты в условиях теплого, а тем более жаркого климата очень недолговечны и быстро разлагаются. Представители названного семейства заметно тяготеют к территориям с прохладным или даже холодным климатом – это средние и высокие (арктические) широты, а также горы. Например, на Тянь‑Шане сплахновые произрастают только высоко в горах, чаще выше верхней границы леса.
Представители семейства Splachnaceae не селятся на субстратах без разбора. Можно наблюдать, как определенные виды явно предпочитают одни субстраты и избегают или, во всяком случае, не селятся на других. Как сказал Кернер [1903]: «Некоторые мхи очень упрямы при выборе субстрата».
Так, наиболее обычные и наиболее распространенные виды рода Splachnum – Sp. rubrum и Sp. luteum – связаны с пометом домашних животных, главным образом коров и лошадей. Потому и встречаются почти исключительно вблизи населенных пунктов. Если же они произрастают на значительном удалении от ближайших сел и выпасов, на расстоянии, например, в несколько десятков километров от них, в малонаселенных горных местностях, то почти всегда оказывается, что где‑то совсем рядом проходит вьючная тропа. Значит, есть конский навоз. В Арктике оба вида чаще всего связаны с пометом северного оленя.
Довольно обычный в высокогорьях Саян и других горных систем Южной Сибири Tetraplodon urceolatus произрастает главным образом на помете северного оленя – и дикого, и домашнего, а в горах Европы – на помете серн, коз и овец.
Два других вида из этого же рода – Tetraplodon angustatus и Tetraplodon mnioides – менее требовательны к характеру субстрата и поэтому наиболее широко распространены. Они встречаются на помете медведя и других лесных животных (в основном хищных), на погадках, экскрементах человека.
В Гималаях сплахновые произрастают большей частью на высотах более 3500 метров над уровнем моря. Часть видов, например, такие как Tetraplodon mnioides и T. angustatus, а также Tayloria nepalensis, приурочены главным образом к помету яков.
Замечательна экология эндемичного для Альп вида Tayloria rudolphiana. Вот что пишет о нем уже цитированный нами Кернер [1903, с. 115]: «Обыкновенно он растет на ветвях старых дерев, особенно старых кленов, в субальпийской полосе, и можно было бы подумать, что он относительно своего субстрата образует исключение между другими Splachnaceae. По если присмотреться ближе, то мы убедимся, что и этот мох живет только на разлагающихся испражнениях животных. Именно, мы всегда находим в субстрате остатки измельченных мышиных и птичьих костей, и не может подлежать сомнению, что эта Tayloria избрала местом поселения испражнения, отложенные хищными птицами на ветвях старых деревьев».
Площадь, занимаемая субстратом, на котором поселяются представители семейства Splachnaceae, ничтожна. Для того чтобы не промахнуться, эти растения или должны производить совершенно баснословное количество спор в надежде, что хотя бы совсем крохотная часть их попадет в нужное место, в нужные условия, или выработать какой‑то механизм, который обеспечивал бы умеренному количеству спор возможность «точного попадания». Ясно, что путь «прицельной стрельбы» экономически гораздо более выгоден. Да и к тому же более надежен. Неудивительно, что Splachnaceae пошли именно по нему.
Но как это осуществляется? Каким образом споры могут попасть с одной кучи помета на другую «прицельно», не разлетаясь вокруг?
Происходит все точно так же, как и перенос пыльцы у цветковых растений – с помощью чужого труда, труда насекомых. Возле разлагающихся животных остатков всегда много насекомых, в том числе навозных мух. Посещая эти участки, насекомые не могут миновать и растущие на них мхи. Тем более, что последние для привлечения насекомых украшают себя чрезвычайно эффектными коробочками с крупной (до 1 сантиметра в диаметре) расширенной нижней частью (гипофизой), окрашенной в яркие красивые тона – желтый, пурпурно‑красный или фиолетовый.
Для большей заметности коробочки помещены на очень высокие ножки и значительно приподняты над поверхностью почвы. Ножка Splachnum luteum, например, может достигать в высоту 15 сантиметров. Это рекорд среди мохообразных!
Но это еще не все. Для полной гарантии посещения насекомыми коробочки некоторые представители семейства издают различные запахи. Так, коробочки Splachnum ampullaceum пахнут молочной кислотой, S. luteum – бутиловыми соединениями.
А дальше все ясно. Перелетая с одной кучки помета (или с одной погадки) на другую, насекомые разносят споры сплахновых. Споры эти, как и надо было ожидать, клейкие и пристают к лапкам насекомых. Так и происходит то самое «точное попадание» спор в нужное место, о котором говорилось выше.
Кстати, Splachnaceae – единственное во всем отделе мохообразных семейство, прибегающее для распространения спор к помощи насекомых. Все остальные надеются главным образом на ветер, точнее, на воздушные течения.
Поселяясь на животных остатках, сплахновые способствуют их скорейшему разложению и тем самым выполняют полезные санитарные функции. Представители этого семейства считаются полусапрофитами. Ну, на сколько процентов они сапрофиты, а на сколько автотрофы, как все остальные зеленые растения, сказать, конечно, нелегко. Но во всяком случае несомненно, что среди мхов, где частичный сапрофитизм сравнительно нередок, сплахновые – в первых рядах. Более сапрофитными, чем они, являются, может быть, еще виды рода Buxbaumia, часто растущие на гниющей древесине.
Это среди мхов. Но вообще среди мохообразных есть один вид, питающийся практически полностью сапрофитно. Это слоевищный печеночник Cryptothallus mirabilis – единственный среди мохообразных вид, лишенный хлорофилла. Растет он в лесах и на болотах под слоем мхов (наиболее часто это различные виды сфагнов) или подстилки из отмирающих растительных остатков, на кислых субстратах. Свет сюда почти не проникает, и фотосинтез невозможен.
Но всем этим разнообразие используемых мохообразными субстратов не исчерпывается. Бриофиты живут еще в дуплах деревьев и кустарников, на старых бетонных сооружениях, включая развалины военных укреплений, на деревянных крышах домов. И так далее.
А одна группа мхов (их около 20 видов), кажется, без всяких кавычек может быть названа бессубстратной. Это те виды, которые могут отрываться от субстрата и свободно переноситься ветром. Обычно это виды, имеющие подушковидную форму. Встречаются они главным образом на открытых местах. Оторвавшись от субстрата, такая подушечка из‑за меняющейся ориентации в пространстве получает возможность расти во все стороны и со временем становится более‑менее сферической. Поэтому бессубстратные мхи чаще всего называют моховыми шариками. В общем это нечто вроде перекати‑поля. Только перекати‑поле отрывается от субстрата и пускается в свободное путешествие в конце вегетационного сезона, когда растение подсыхает и отмирает. Путешествует отмершее растение. А моховой шарик живой. Внутри него бывает небольшой камешек, вместе с которым он когда‑то оторвался от субстрата, или песок. Нередко центральная часть шарика отмершая. Так что какой‑то минимум питательных веществ шарик перетаскивает на себе или, точнее, внутри себя. А влагу получает из атмосферы. Или из того участка субстрата, на котором он временно остановился. Таким участком может быть и поверхность ледника! Так как мох темнее льда, он больше нагревается, лед под ним слегка протаивает, и мох оказывается в небольшом углублении. При этом он, понятно, увлажняется.
Моховые шарики могут образовывать виды, растущие, как уже говорилось, главным образом на открытых местах. Но образуют их в виды, растущие в лесах. Отмечены такие моховые шарики в Арктике, в Субарктике и Субантарктике, в умеренных широтах обоих полушарий. Другими словами, распространены они довольно широко.
Обычные размеры моховых шариков 6–9 сантиметров в диаметре (или в длину, есть форма шарика не вполне сферическая). Максимальный размер около 15 сантиметров.
Кроме моховых шариков «свободный образ жизни» (без прикрепления к субстрату) могут вести еще несколько видов лишайников, в том числе и знаменитая «манна небесная». Все остальные растения всю жизнь прикованы к одному месту. Путешествовать могут или их отмершие части, или семена и споры.
Вот так в кратком изложении выглядит географическое распространение и экологическое распределение мохообразных. О насекомых в начале главы мы упомянули не случайно – по способности проникать в любую «щель», по необозримости географического распространения и необъятности экологических условий, при которых могут жить мохообразные, их вполне можно сравнить именно с насекомыми. Конечно, такое сравнение условно уже хотя бы потому, что насекомые – животные и многие стороны их жизни регулируются другими законами. По зато такое сравнение дает представление о масштабах географического распространения мохообразных, объясняя вместе с тем, почему при столь широком распространении мохообразные в общем малозаметны.
Дата добавления: 2016-01-26; просмотров: 859;