Окислительно‑восстановительная система … Окислительный потенциал
ДПН/ДПН∙2Н … ‑ 0,32
ФАД/ФАД‑ 2Н … ‑ 0,22
b(Fe3+)/b(Fe2+)* … ‑ 0,05
c(Fe3+)/c(Fe2+)* … + 0,25
a(Fe3+)/aFe2+)* … + 0,29
a3(Fe3+)/a3(Fe2+* … + 0,30
* цитохромы
Как я уже говорил, при дегидрогенизации вещества уровень свободной энергии снижается на 35–70 килокалорий на моль вещества, но организм не может использовать высвобождающуюся энергию, если только не запасет ее в виде высокоэнергетических фосфатных связей. Насколько известно, в ходе одной реакции может образоваться только одна высокоэнергетическая связь. Поскольку запасти таким образом можно только 8 килокалорий на моль вещества, то перевод одной реакции дегидрогенизации в одну реакцию образования высокоэнергетической фосфатной связи означал бы потерю семи восьмых энергии.
Наличие длинной дыхательной цепочки позволяет живой ткани разбить процесс уменьшения свободной энергии на фрагменты, на каждом из которых можно создавать по одной высокоэнергетической фосфатной связи. В начале 50‑х годов XX века достаточно убедительно было показано, что таким образом в среднем на основе одной реакции дегидрогенизации удается создать по три высокоэнергетических фосфатных связи. Таким образом, эффективность использования энергии достигает хотя бы 35 процентов.
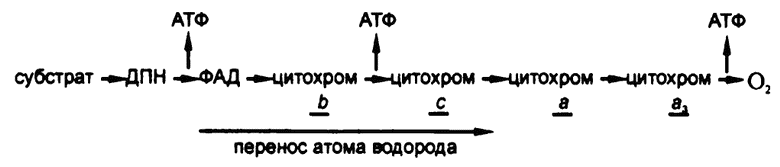
Рис. 63. Окислительное фосфорилирование
Естественно, возникает вопрос, а где же именно, на каком участке дыхательной цепочки формируются высокоэнергетические фосфатные связи. Для ответа на этот вопрос попытаемся действовать методом исключения.
Система с участием дегидрогеназы молочной кислоты производит три высокоэнергетические фосфатные связи на каждую потребляемую молекулу молочной кислоты. Однако система с участием дегидрогеназы янтарной кислоты произведет лишь две высокоэнергетические фосфатные связи на каждую потребляемую молекулу янтарной кислоты. Дегид‑рогеназа янтарной кислоты – флавофермент, так что при дегидрогенизации янтарной кислоты из процесса выпадает этап пиридин‑фермента. Поскольку этому выпадению соответствует потеря одной реакции образования высокоэнергетической фосфатной связи, то остается сделать вывод, что одна такая связь образуется именно при переходе от ДПН к ФАД.
Путем добавления в дыхательную систему различных химических веществ можно исключать из цепочки те или иные звенья и отмечать, сократится ли при этом количество образуемых высокоэнергетических фосфатных связей. Итоговые предположения об участках образования этих связей приведены на рис. 63.
Каковы же механизмы реакций «окислительного фосфорилирования» (то есть производства высокоэнергетических фосфатных связей путем окисления, представленного в виде переноса атомов водорода)? Это до сих пор неизвестно. Если излагаемая в данной книге информация производит впечатление, что наука сумела решить все проблемы, то заверяю вас – это впечатление ошибочно.

Рис. 64. АТФ и цикл Кребса
Без знания точных подробностей производства высокоэнергетических связей тем не менее возможно подсчитать количество производимого на каждой стадии катаболизма молочной кислоты – результаты приведены на рис. 64.
Как видите, результатом катаболизма молочной кислоты на углекислоту и воду, в отношении которого калориметр показывает цифру в 325 калорий на моль вещества[9], являются 18 молекул АТФ. Если считать, что при формировании каждой высокоэнергетической фосфатной связи уровень свободной энергии возрастает на 8 килокалорий на моль вещества, то в целом в результате катаболизма каждого моля молочной кислоты организм переводит в высокоэнергетические фосфатные связи и запасает 144 килокалории химической энергии. 144 из 325 – это около 45 процентов, в общем, неплохая эффективность.
В какой же части клетки проходят процессы окислительного фосфорилирования? В данной книге я еще ни разу не предпринимал попыток описать устройство клетки. А клетка устроена очень сложно, она обладает множеством специализированных частей (органелл) со специализированными свойствами, и чем более тонкие инструменты оказываются в руках цитологов, тем более тонкие свойства органелл удается обнаружить.
Начнем с того, что каждая неповрежденная клетка имеет внутреннюю часть небольшого объема, которая носит название «клеточное ядро», тонкой мембраной отделенное от остальной части клетки (цитоплазмы). Главная задача ядра – обеспечение размножения клеток и точный перенос генов, управляющих химическими свойствами, от материнской клетки к дочерней. Однако к процессу производства энергии ядро никакого отношения не имеет и ни флавоферментов, ни цитохром не содержит. Для дегидрогенизации как таковой не требуется ничего, кроме пиридин‑ферментов, которые сами по себе кислород не используют. Иными словами, ядро – система анаэробная, так что искать место проведения энергопроизводящих реакций с потреблением кислорода следует в цитоплазме.
Различные органеллы цитоплазмы (да и ядра) реагируют на разные красители по‑разному – одни поглощают, другие нет, так что их можно «помечать» с помощью соответствующих красителей, специфичных для каждой из органелл. Тогда клеточные частицы, обычно неразличимые глазом, начинают сверкать всеми цветами радуги.
В 1898 году немецкий цитолог С. Бенда с помощью комплекса красителей сумел обнаружить, что небольшие гранулы в цитоплазме огромного множества клеток окрашиваются подозрительно контрастно. Он назвал эти гранулы «митохондриями» (от греческого «хрящевые нити»). Название было неудачным, поскольку к хрящам эти нити никакого отношения не имеют. Тем не менее название это прижилось, как и многие другие ошибочные имена, и мы пользуемся им по сей день.
На протяжении примерно одного поколения функции митохондрий оставались неизученными, и они были для всех просто «еще одним видом клеточных частиц». Однако в 30‑х годах XX века с развитием ультрацентрифуг (см. главу 16) стало возможным разложение клетки на составляющие путем ее разрушения и осаждения по очереди различных содержащихся в ней веществ. Таким образом были получены и суспензии, состоящие из одних митохондрий.
А в 40‑х годах XX века появились и новые микроскопы с небывалой увеличительной мощностью. Теперь вместо лучей света, фокусируемых с помощью оптических линз, стали использоваться пучки электронов, фокусируемые с помощью магнитных полей. С помощью таких электронных микроскопов наконец‑то стало возможным как следует рассмотреть митохондрии, диаметр которых составляет всего один‑три микрона, то есть в тысячу раз меньше самой клетки.
Использование электронного микроскопа позволило установить, что митохондрия окружена двойной мембраной, рассекающей тело органеллы, разделяющей ее на части и многократно увеличивающей, таким образом, площадь поверхности митохондрии.
В ходе экспериментов выяснилось, что суспензия митохондрии катализирует все без исключения реакции, входящие в цикл Кребса. Так стало ясно, что «электростанцией» клетки служит именно митохондрия, мембраны которой представляют собой скопление ферментов и коферментов, необходимых для катаболизма пищевых веществ и производства высокоэнергетических фосфатных связей. Было подсчитано, что в среднем отдельная митохондрия представляет собой около 10 000 «сборочных пунктов», на каждом из которых может происходить полный цикл катаболизма, отщепляться атом водорода и создаваться высокоэнергетическая фосфатная связь.
Более подробная информация о молекулярном устройстве митохондрии наверняка еще появится по мере разработки более мощных научных инструментов.
Глава 24.
Дата добавления: 2016-01-26; просмотров: 1122;