Обезжиренное молоко
 |
Подкисление до рН 4,6
температура 20оС


Осадок – казеиновая Фильтрат – сывороточ-
фракция (78-85 %) ные белки (15-22 %)

Нагревание до 95оС,
в течение 20 мин.,
 фильтрование
фильтрование


Осадок – фракция Фильтрат – фракция
термолабильных термостабильных
сывороточных белков сывороточных белков
 | |
Полунасыщение Добавление 12%-
(NH4)2SO4 или ной трихлорук-
насыщение MgSO4 сусной кислоты
 | |


 Осадок- Фильтрат - Осадок - Фильтрат –
Осадок- Фильтрат - Осадок - Фильтрат –
лактогло- лактальбумино- протеозо- небелковые
булиновая вая фракция, пептоны азотистые
фракция растворимая в соединения
(иммуно- растворах солей
глобули- (основные сыво-
ны) роточные белки)
 |  |  |  | ||||

 ИrG1 (1,2-3,3%) b-лактоглобулин Электрофо- мочевина
ИrG1 (1,2-3,3%) b-лактоглобулин Электрофо- мочевина

 ИrG2 (0,2-0,7%) (АВСDEFG) (7-12%) ретические свободные
ИrG2 (0,2-0,7%) (АВСDEFG) (7-12%) ретические свободные
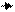
 ИrM (0,2-0,7%) a-лактальбумин фракции - аминокис-
ИrM (0,2-0,7%) a-лактальбумин фракции - аминокис-

 ИrA (0,1-0,7%) (АВ) (2-5%) 3,5,8 быст- лоты
ИrA (0,1-0,7%) (АВ) (2-5%) 3,5,8 быст- лоты

 альбумин сыворот- рый и 8 - креатин
альбумин сыворот- рый и 8 - креатин

 ки крови (0,7-1,2%) медленный креатинин
ки крови (0,7-1,2%) медленный креатинин
 (2-6%) аммиак
(2-6%) аммиак
Рис. 5.2. Фракционирование сывороточных белков
Ca3(PO4)2, а часть образует комплекс с a-лактальбумином и с c-казеином мицелл казеина и осаждается вместе с ними при кислотной и сычужной коагуляциях казеина. Образование комплекса β-лактоглобулин-α-лактальбумин-c-казеин значительно ухудшает атаку χ-казеина сычужным ферментом и снижает термоустойчивость мицелл казеина.
a-Лактальбумин. В количественном отношении занимает второе место после b-лактоглобулина в содержании сывороточных белков (от 20 до 25%). Представляет собой компактный глобулярный белок, частицы которого имеют самые малые размеры по сравнению с частицами других белков молока.
a-Лактальбумин является гетерогенным белком (содержит главный компонент, имеет два генетических варианта, а также минорные компоненты, некоторые из которых являются гликопротеидами). В молоке a-лактальбумин тонкодиспергирован, не коагулирует в изоэлектрической точке в силу своей большой гидратированности, не свертывается под действием сычужного фермента, относительно термостабилен. Повышенная устойчивость его к нагреванию обусловлена наличием в его молекуле большого количества дисульфидных связей.
Альбумин сыворотки крови – это относительно крупный глобулярный белок, его полипептидная цепь состоит из 582 аминокислотных остатков, содержит 17 внутримолекулярных дисульфидных связей и только одну свободную сульфгидрильную группу, термолабилен.
Альбумин сыворотки крови не синтезируется в молочной железе, а поступает в молоко из крови. Содержание этой фракции в молоке увеличивается при нарушении функций молочной железы (при заболеваниях маститом).
Лактоглобулиновая фракция сывороточных белков включает в себя термолабильные белки, которые осаждаются из молочной сыворотки при полунасыщении сульфатом аммония или насыщении сульфатом магния. К б елкам этой фракции относятся иммуноглобулины. В нормальном молоке массовая доля иммуноглобулинов составляет от 1,9 до 3,3% общего содержания белков, а в молозиве они составляют около 90% всех сывороточных белков.
Иммуноглобулины – это высокомолекулярные белки (гликопротеиды), выполняющие функцию антител. Антитела – вещества, образующиеся в организме при введении в него различных чужеродных белков (антигенов) и нейтрализующие их действие. Иммуноглобулины молока обладают резко выраженными свойствами агглютинации – склеивания микробов и других чужеродных клеток, а также шариков жира. Из выделенных основных групп иммуноглобулинов: G, A и М в молоке преобладают иммуноглобулины ИrG1 (1,2-3,3 % общего содержания белков).
Термостабильная фракция сывороточных белков включает в себя протеозо-пептонную фракцию, осаждаемую трихлоруксусной кислотой из фильтрата сыворотки после осаждения термолабильных сывороточных белков. Доля протеозо-пептонной фракции составляет 2-6% всех белков молока. Эта фракция неоднородна по составу, состоит из четырех компонентов, которые обозначают 3, 5, 8 «быстрый» и 8 «медленный» (по скорости передвижения в электрическом поле).
Компонент 3 имеет высокое содержание углеводов, а остальные являются фосфопептидами, образующимися вместе с g-казеинами при гидролизе b-казеина протеиназами молока. Содержание протеозопептонной фракции увеличивается в процессе длительного хранения молока при 3-5 оС. Кроме этого, по содержанию этой фракции судят о глубине протеолитических процессов, например, при созревании сыров.
Кроме вышеназванных белков в молоке содержатся другие белки и вещества белковой природы, концентрация которых выражается в миллиграммах на 100 г молока. К ним относятся белки оболочек жировых шариков, лактоферрин, белки, обладающие антибактериальными и ферментативными свойствами.
Белки оболочек жировых шариков. Содержание их составляет около 1% всех белков молока. Отличаются от казеина повышенным содержанием цистина и меньшим содержанием кислых аминокислот. Это, как правило, гликопротеиды, содержащие от 15 до 50% углеводов (галактозу, N-ацетилгалактозамин, N-ацетилглюкоза-мин, N-ацетилнейраминовую кислоту), образующие с лецитином липопротеидный комплекс – структурный элемент оболочек жировых шариков, способствующий их стабильности во время технологической обработки молока.
Лактоферрин представляет собой гликопротеид, содержащий два атома железа, обладающий антибиотической активностью по отношению к бактериям группы кишечной палочки, связывая ионы железа и делая их недоступными для бактериальных клеток. Лактоферрин усиливает бактерицидное действие лизоцима. Содержание его в молоке составляет не более 30 мг на 100 г , а в молозиве – в 10-15 раз больше. В организме новорожденного лактоферрин выполняет транспортную функцию – связывает и переносит ионы железа.
Лизоцим – компонент белковой природы с ферментативными свойствам, вызывающий лизис бактериальных клеток (нарушение клеточной оболочки).
Низин– полипептид, обладающий антибактериальными свойствами по отношению к споровым бактериям. Низин продуцируют некоторые виды микрофлоры молока и бактериальных заквасок.
Ферменты – вещества белковой природы, биологическая функция которых состоит в ускорении (катализе) течения химических реакций в биологических системах. Все эти вещества в одних случаях – простые белки, а в других – сложные, содержащие в своем составе небелковые компоненты – коферменты. В молоке содержится более 20нативных ферментов, а также присутствуют многочисленные внеклеточные и внутриклеточные ферменты, продуцируемые микрофлорой молока и бактериальных заквасок.
Структура белков
Белковые молекулы содержат от 100 до нескольких тысяч аминокислот в пептидных цепях, упорядоченных в пространстве. Для характеристики строения белков введены понятия о первичной, вторичной и третичной структуре. И если речь идет о белках, образующих мицеллы, - о четвертичной структуре. В настоящее время изучена первичная структура большинства компонентов казеина, b-лактоглобулина, a-лактальбумина, протеозо-пептонов. А также получены некоторые сведения о вторичной, третичной и четвертичной структурах основных белков молока.
Первичная структура определяется числом и расположением a-аминокислот, конфигурацией связей в полипептидных цепях и, если белки состоят из нескольких полипептидных цепей, - местоположением и типом поперечных связей. Для выяснения первичной структуры белков необходимо знать их аминокислотный состав. Аминокислотный состав белков молока в настоящее время достаточно изучен. Намного труднее установить расположение аминокислот в полипептидных цепях. Многообразие белков, в том числе молочных, обусловлено различиями расположения 20 составляющих их аминокислот (последовательностью расположения аминокислот). При исследовании первичной структуры белка важно определить концевые группы полипептидных цепей. Каждая полипептидная цепь имеет концевую аминогруппу и концевую карбоксильную группу:
Н2N ¾ СН ¾ LLLL ¾ СН ¾ СООН
R R
 На различной способности концевых групп реагировать с химическими веществами, основаны методы их идентификации. Последовательное отщепление аминокислот от полипептидной цепи позволяет установить последовательность расположения аминокислот, то есть первичную структуру белка. Представление о сложной первичной структуре белков молока дает, например, схема последовательности аминокислот во фракциях казеина: aS1 – 199 аминокислот, b-казеин – 209; c-казеин – 169. Аминокислоты располагаются таким образом, что конец одной аминокислоты связан с началом другой пептидной связью. Пептидная связь образуется в результате выделения молекулы воды из карбоксильной группы одной аминокислоты и аминогруппы другой.
На различной способности концевых групп реагировать с химическими веществами, основаны методы их идентификации. Последовательное отщепление аминокислот от полипептидной цепи позволяет установить последовательность расположения аминокислот, то есть первичную структуру белка. Представление о сложной первичной структуре белков молока дает, например, схема последовательности аминокислот во фракциях казеина: aS1 – 199 аминокислот, b-казеин – 209; c-казеин – 169. Аминокислоты располагаются таким образом, что конец одной аминокислоты связан с началом другой пептидной связью. Пептидная связь образуется в результате выделения молекулы воды из карбоксильной группы одной аминокислоты и аминогруппы другой.
¾ С ¾ NH ¾ - пептидная связь;
О
R
— 
 HC —NH — C — CH — NH — C — CH — NH — C — ΛΛ… -
HC —NH — C — CH — NH — C — CH — NH — C — ΛΛ… -
R O O R O
- пептидные связи в полипептидной цепи
Пептидные связи очень прочные (валентные).
Многие белки имеют несколько полипептидных цепей, соединенных друг с другом дисульфидными связями R – S – S – R цистина, причем оба конца молекулы цистина входят в состав двух различных цепей. Эти дисульфидные связи могут находиться и внутри полипептидной цепи, свернутой в виде спирали. Кроме этого, между фосфорной кислотой и серином в молекуле белка возможно образование фосфатноэфирной связи (рис. 5.3).
Рис. 5.3. Типы связей первичной структуры белков:
1 – пептидная связь; 2 – дисульфидная связь;
3 – фосфатноэфирная связь
О‾
– Н2С – О – Р – О – СН2 – фосфатноэфирная связь
О
Эта связь непрочная, при нагревании молока она разрушается. Таким образом, первичная структура белков основана на главных валентных связях: пептидных и дисульфидных. Они стабильны, не разрушаются при обработке и переработке молока (при энергетических воздействиях). Первичная структура белков молока разрушается только при ферментативном гидролизе белка (например, в процессе длительного хранения молока или при созревании сыров), либо при кислотном гидролизе белка под действием концентрированной соляной кислоты (используется в исследовательских целях при определении аминокислотного состава белков).
 Вторичная структура белков отражает порядок пространственной ориентации полипептидной цепи – конформацию, зависящую от взаимного расположения аминокислотных остатков полипептидной цепи. Полипептидные цепи белковых молекул могут иметь спиралевидную или зигзагообразную укладку, стабилизированую за счет водородных связей: С = О….Н – N
Вторичная структура белков отражает порядок пространственной ориентации полипептидной цепи – конформацию, зависящую от взаимного расположения аминокислотных остатков полипептидной цепи. Полипептидные цепи белковых молекул могут иметь спиралевидную или зигзагообразную укладку, стабилизированую за счет водородных связей: С = О….Н – N
N ─ H ─ ─ ─ ─ ─ O ═ C N ─ H
R – C – H H – C – R H – C – R
C = O ─ ─ ─ ─ ─ H —N C = O
 Н – N C = O ─ ─ ─ ─ ─ H —N
Н – N C = O ─ ─ ─ ─ ─ H —N
R ─ C ─ H R ─ C ─ H H ─C ─ R

Водородные мостики между
полипептидными цепями
Водородные связи могут образовываться также внутри полипептидной цепи. Если вообразить, что полипептидная цепь намотана на цилиндр, то получится спиралевидная конфигурация, в которой атомы кислорода карбонильной группы и водород амидогруппы приближены друг к другу на такое расстояние, при котором образуются водородные связи. Водородные связи объясняются полярным характером и остаточными валентностями присутствующих атомных группировок. Если отрицательно заряженный атом кислорода, то есть атом кислорода со свободной парой электронов, приближается к поляризованному атому водорода амидогруппы, то в этой случае действуют силы электростатического взаимодействия.
Спиралевидная конформация полипептидных цепей получила название α-спирали. На каждый виток спирали приходится 3,7 аминокислотного остатка. Все боковые цепи аминокислотных остатков расположены вне спирали. Значительные расстояния между боковыми радикалами исключают возможность их взаимодействия в составе одной α-спирали. В белках вероятна и возможность существования других структур (не только конформация α-спираль). Другая конформация – β-складчатая структура (зигзагообразная) – представляет систему параллельно или антипараллельно расположенных участков одной или нескольких полипептидных цепей, соединенных водородными связями. В отличие от α-спирали водородные связи в такой системе перпендикулярны направлению полипептидных цепей.
В настоящее время доказано, что αS1-, β- и κ-казеины имеют незначительное количество α-спиральных участков (от 1 до 6% и более по некоторым данным), то есть молекулы казеинов имеют малоупорядоченную структуру. β-Лактоглобулин и α-лактальбумин содержат от 10 до 26% α-спиральных участков, а остальное составляет β-складчатая структура (β-конформация) и неупорядоченная структура.
Кроме водородных связей вторичную структуру (пространственную ориентацию полипептидных цепей) обусловливают дисульфидные связи: R – S – S – R.
Водородная связь обладает незначительной энергией, ее нельзя считать истинной химической связью. Она может расщепляться при обработке и переработке молока, например при высокотемпературной пастеризации. Разрыв водородных связей внутри спирали или β складчатой конформации полипептидной цепи ведет к ее развертыванию, а освободившиеся побочные валентности могут вступать во взаимодействие с другими цепями.
Третичная структура отражает способ укладки полипептидных цепей в глобулярных белках с образованием компактной структуры. Формирование третичной структуры обусловлено тем, что аминокислоты пролин и оксипролин своей плоской конфигурацией нарушают α-спираль, вследствие чего происходит ее развертывание и образование компактных шарообразных структур.
Стабильность третичной структуры обусловлена силами взаимодействия полярных и неполярных связей боковых цепей, а также водородными связями. Силы, исходящие от неполярных боковых цепей, представляют собой энергию гидрофобных связей
─ С – R ◠◠◠ R – C ─
(силы молекулярного притяжения Ван-дер-Ваальса-Лондона). Взаимодействие между полярными участками боковых цепей стабилизируется ионной связью
— С – N+H3 ~ ‾OOC – C —
Боковые цепи, обеспечивающие сохранение третичной структуры, и функциональные группы находятся внутри шарообразных конфигураций. Остаточные функциональные группы на поверхности частиц предопределяют химические и физические свойства белковых молекул, причем от ионизированных карбоксильных и аминогрупп зависят растворимость и стабильность белков. В форме ионов с электрическим зарядом они присоединяют воду, то есть гидратируются. Гидратная воды образует защитную оболочку вокруг белковых молекул в коллоидном растворе. У каждого вида белка своя структура. Она формируется при биосинтезе белка в организме. Если связи, обусловливающие конформацию (то есть вторичную и третичную структуру) расщепляются, то это ведет к изменению структуры белка. Причем к необратимому изменению, после устранения факторов, вызывающих расщепление, первоначальная структура белка не восстанавливается, вследствие чего изменяются его свойства. В таком случае белок считается денатурированным.
Четвертичная структура. Казеины, как некоторые другие белки, могут образовывать непрочно связанные друг с другом мицеллы, то есть формировать четвертичную структуру. Мицеллы – это агрегаты частиц, состоящие из субединиц (субмицелл), которые легко разрушаются под воздействием внешних факторов. Связь между субмицеллами в мицеллах осуществляется, по-видимому, через кальций-фосфатные мостики, так как величина казеиновых частиц в значительной степени зависит от содержания кальция в молоке. После удаления кальция мицеллы казеина распадаются на субмицеллы.
Как уже отмечалось в разделе 5.1 все фракции казеина являются фосфопротеидами и в присутствии кальция, цитратов и фосфатов склонны самоассоциироваться и взаимодействовать друг с другом с образованием ассоциатов различных размеров. По-видимому между фракциями казеина могут возникать связи различных типов: гидрофобные взаимодействия, ионные, водородные и дисульфидные. Способность казеинов к ассоциации зависит от температуры, рН и ионной силы молока. Установлено, что казеины содержатся в молоке в виде растворимых казеинатов кальция в сочетании с коллоидным фосфатом кальция в виде казеинаткальцийфосфатного комплекса (ККФК).
В настоящее время с помощью электронномикроскопических исследований установлено, что ККФК образует мицеллы, которые имеют сферическую форму с диаметром от 40 до 300 нм (средний диаметр составляет около 100 нм) и являются высокоорганизованными структурными единицами со средней молекулярной массой 6.108.
Под четвертичной структурой белка понимают структуру мицелл казеина. Несмотря на многочисленность исследований в этом направлении, вопрос о модели структуры казеина все еще остается дискуссионным. К настоящему времени известны более десяти моделей структуры мицелл казеина, которые предполагают сцепление в мицелле, как отдельных полипептидных цепей, так и субмицелл между собой, в основном, с помощью коллоидного фосфата кальция. В основе большинства моделей лежит субмицеллярный принцип построения мицеллы, то есть субмицеллы рассматриваются как структурные субъединицы мицелл. Сведения о характере комбинаций фракций казеина в субмицеллах достаточно противоречивы. Эти противоречия касаются соотношения фракций казеина в субмицеллах, относительного распределения их в объеме субмицелл (гидрофобное ядро и гидрофильная поверхность), зависимости соотношения фракций казеина от размеров субмицелл.
Первой моделью субмицеллярного строения мицелл является модель, разработанная Мором. Другие модели этого типа можно рассматривать как ее модификации, в том числе и модель, разработанную Д.Г.Шмидтом (рис.5.4).
Автор предполагает, что мицелла казеина состоит из субмицелл диаметром 10-20 нм. Субмицеллы состоят из αS1-, αS2-, β- и κ -казеинов в соотношении 3:1:3:1. Фракции казеина связаны фосфатом кальция и в субмицелле располагаются таким образом, что гидрофобные участки фракций находятся внутри ядра, а гидрофильные участки κ -казеина и фосфатные группы αS1-, αS2-и β-казеинов – на поверхности.
Субмицеллы объединяются в мицеллы с помощью коллолидного фосфата кальция (на рисунке обозначен точками). Ряд исследователей не исключают возможность мицеллообразования казеина за счет гидрофобных взаимодействий между субмицеллами.
Субмицеллы с незначительным содержанием κ -казеина или без него расположены внутри мицеллы, а с высоким содержанием κ -казеина – на ее поверхности. Таким образом, поверхность мицеллы казеина за счет электрических зарядов связывает значительное количество воды, что обусловлено полярностью ее молекул. На образовавшемся слое адсорбируются другие частицы воды. По мере утолщения слоя связанной воды новые молекулы воды все слабее удерживаются коллоидной частицей.
С появлением новых и развитием традиционных методов исследований структура модели мицелл казеина уточнялась и совершенствовалась. С помощью элетронномикроскопических исследований удалось установить, что на поверхности мицелл имеется «волосковый слой», представляющий собой выступающие с поверхности
мицелл гидрофильные участки полипептидных цепей χ-казеина – гликомакропептиды. «Волосковый слой» распространяется в дисперсионную среду на 10-15 нм и препятствует тесному сближению мицелл.
Рис. 5.4. Модель субмицеллы (а) и мицеллы казеина (б)
Обобщенная субмицеллярная модель мицеллы казеина представлена схематически на рис. 5.5.
Таким образом, строение белков молока следующее. Белки состоят из крупных молекул (полипептидных цепей) с относительной молекулярной массой свыше 10000, способных агрегировать друг с другом в субмицеллы и мицеллы. Свойства белков определяются количеством, видом и последовательностью расположения аминокислот (т.е. первичной структурой), их пространственной ориентацией и способом укладки полипептидных цепей (вторичной и третичной структурой), а также взаимодействием агрегатов основных фракций субмицелл (из αS1-, αS2-, β- и χ –казеинов) в мицеллах (четвертичной структурой). Силы, обусловливающие ту или иную структуру отличаются по своей природе и прочности.
Рис. 5.5. Субмицеллярная модель мицеллы казеина
5.3. Состав белков: элементарный и аминокислотный
Белки молока содержат все органические природные вещества: углерод, водород, кислород и характерный для белков элемент – азот.
Кроме этого в их состав входят сера и фосфор.
Для характеристики белков служит прежде всего наличие в них азота, серы и фосфора. На результатах элементарного анализа, в частности азота, основаны методы определения белков в молоке. Для определения содержания азота в органических соединениях был разработан метод Кьельдаля (в 1883 году), который и в настоящее время служит надежным способом определения белков в молоке и молочных продуктах. Количество белков устанавливается по содержанию азота в анализируемой пробе молока с учетом коэффициента перевода. Путем сжигания пробы и последующей фиксации азота определяют его содержание. Коэффициент пересчета принимают равным 6,38, исходя из того, что в 100 г чистого казеина содержится 15,65% азота (по результатам анализа элементарного состава казеина – (100:15,65=6,38). Средний элементарный состав казеина в %:азота – 15,65; углерода – 53,00;кислорода – 22,60; водорода – 7,10; серы – 0,80; фосфора – 0,85).
Элементарные составы казеина и сывороточных белков значительно различаются содержанием фосфора. По содержанию фосфора различаются между собой и компоненты казеина.
Казеин – это ярко выраженный фосфопротеид, а сывороточные белки либо совсем не содержат фосфора, либо его ничтожно мало.
Фосфор присутствует в форме фосфорнокислых сложных эфиров оксиаминокислот серина и треонина. Тип связи фосфора в казеине играет определенную роль в образовании кальций-фосфатных мостиков в казеиновом комплексе и между субмицеллами казеина:
ОН
- СН2 – О – Р = О - моноэфирная связь;
ОН
ОН
- СН2 – О – Р – О – СН2 – - диэфирная связь
О
ОН ОН
- СН2 – О – Р – О – Р – О – СН2 – - пирофосфатэфирная связь
О О
В сывороточных белках серы больше, чем в казеинах. β-Лактоглобулин содержит SH-группы и может действовать как антиокислитель. Наличие серы обусловлено присутствием в сывороточных белках серосодержащих аминокислот – метионина, цистина и цистеина. Они оказывают влияние на изменения вкуса молока при тепловой обработке. Большую роль в определении свойств белков, их структуры играет их аминокислотный состав.
Знание аминокислотного состава белков позволяет установить последовательность расположения аминокислот в молекуле белка и выявить его структуру. По количеству и видам аминокислот можно судить о наличии функциональных групп, то есть о их возможных реакциях, а от содержания незаменимых аминокислот зависит питательная и биологическая ценность белков.
Белки молока содержат почти все аминокислоты, обычно встречающиеся в белках: алифатические, оксиаминокислоты, дикарбоновые, диаминокислоты, ароматические, серосодержащие. Основные белки молока по сравнению с другими глобулярными белками пищевых продуктов содержат сравнительно много лейцина, изолейцина, лизина, глутаминовой кислоты. По содержанию и соотношению незаменимых аминокислот белки молока относятся к биологически полноценным белкам.
Сравнение аминокислотного состава основных белков молока позволяет сделать следующие выводы:
- сывороточные белки содержат больше незаменимых аминокислот (изойлецина, лейцина, лизина, метионина, фенилаланина, триптофана, треонина, валина), чем казеины;
- казеины характеризуются более высоким содержанием дикарбоновых аминокислот – глутаминовой и аспарагиновой;
- высокое содержание пролина и оксипролина в казеинах обусловливает преобладание в них складчатой конфигурации полипептидных цепей (эти аминокислоты своей плоской конфигурацией нарушают α-спираль, что ведет к ее развертыванию и образованию компактной шарообразной структуры).
Дата добавления: 2016-01-20; просмотров: 1315;