Ретикулярная формация.
Термин ретикулярная формация предложил в 1865 году немецкий ученый О. Дейтерс. Под этим термином Дейтерс понимал разбросанные в стволе головного мозга клетки, окруженные множеством волокон, идущих в различных направлениях. Именно сетевидное расположение волокон, связывающих между собой нервные клетки, послужило основой для предложенного названия.
В настоящее время морфологами и физиологами накоплен богатый материал о строении и функциях ретикулярной формации. Установлено, что структурные элементы ретикулярной формации локализуются в целом ряде мозговых образований, начиная с промежуточной зоны шейных сегментов спинного мозга (VII пластина), и заканчивая некоторыми структурами промежуточного мозга (интраламинарными ядрами, таламическим ретикулярным ядром). Ретикулярная формация состоит из значительного числа нервных клеток (в ней содержится почти 9/10 клеток всего ствола мозга). Общие черты строения ретикулярных структур - наличие особых ретикулярных нейронов и отличительный характер связей.
Рис. 1. Нейрон ретикулярной формации. Сагиттальный разрез ствола мозга крысенка.
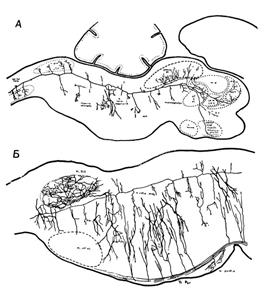
На рисунке А представлен только один нейрон ретикулярной формации. Видно, что аксон разделяется на каудальный и ростральный сегменты, большой протяженности, со множеством коллатералей. Б. Коллатерали. Сагиттальный разрез нижней части ствола мозга крысенка, показывающий соединения коллатералей большого нисходящего пути (пирамидный путь) с ретикулярными нейронами. Коллатерали восходящих путей (сенсорные пути), отсутствующие на рисунке, соединяются с ретикулярными нейронами подобным же образом (по Шейбэлу М. Э. и Шейбэлу А. Б.)
Наряду с многочисленными отдельно лежащими нейронами, различными но форме и величине, в ретикулярной формации головного мозга имеются ядра. Рассеянные нейроны ретикулярной формации прежде всего играют важную роль в обеспечении сегментарных рефлексов, замыкающихся на уровне ствола головного мозга. Они выступают в качестве вставочных нейронов при осуществлении таких рефлекторных актов, как моргание, роговичный рефлекс и т. д.
Выяснено значение многих ядер ретикулярной формации. Так, ядра, расположенные в продолговатом мозге, имеют связи с вегетативными ядрами блуждающего и языкоглоточного нервов, симпатическими ядрами спинного мозга, они участвуют в регуляции сердечной деятельности, дыхания, тонуса сосудов, секреции желез и т. д.
Установлена роль голубого пятна и ядер шва в регуляции сна и бодрствования. Голубое пятно, находится в верхнелатеральной части ромбовидной ямки. Нейроны этого ядра продуцируют биологически активное вещество - норадреналин, который оказывает активирующее воздействие на нейроны вышележащих отделов мозга. Особенно высока активность нейронов голубого пятна во время бодрствования, во время глубокого сна она угасает почти полностью. Ядра шва располагаются по срединной линии продолговатого мозга. Нейроны этих ядер вырабатывают серотонин, который вызывает процессы разлитого торможения и состояние сна.
Ядра Кахаля и Даркшевича, относящиеся к ретикулярной формации среднего мозга, имеют связи, с ядрами III, IV, VI, VIII и XI пар черепных нервов. Они координируют работу этих нервных центров, что очень важно для обеспечения сочетанного поворота головы и глаз. Ретикулярная формация ствола головного мозга имеет важное значение в поддержании тонуса скелетной мускулатуры, посылая тонические импульсы на мотонейроны двигательных ядер черепных нервов и двигательных ядер передних рогов спинного мозга. В процессе эволюции из ретикулярной формации выделились такие самостоятельные образования, как красное ядро, черное вещество.
По структурно-функциональным критериям ретикулярная формация делится на 3 зоны:
1. Медианную, расположенную по средней линии;
2. Медиальную, занимающую медиальные отделы ствола;
3. Латеральную, нейроны которой лежат вблизи сенсорных образований.
Медианная зона представлена элементами шва, состоящие из ядер, нейроны которых синтезируют медиатор – серотонин. Система ядер шва принимает участие в организации агрессивного и полового поведения, в регуляции сна.
Медиальная (осевая) зона состоит из мелких нейронов, которые не ветвятся. В зоне располагается большое количество ядер. Встречаются также крупные мультиполярные нейроны с большим числом густо ветвящихся дендритов. Они образуют восходящие нервные волокна в кору больших полушарий и нисходящие нервные волокна в спинной мозг. Восходящие пути связи медиальной зоны оказывают активирующее влияние (прямо или опосредованно через таламус) на новую кору. Нисходящие пути оказывают тормозящее влияние.
Латеральная зона – к ней относятся ретикулярные образования, расположенные в стволе мозга вблизи сенсорных систем, а также ретикулярные нейроны, лежащие внутри сенсорных образований. Основным компонентом этой зоны является группы ядер, которые примыкают к ядру тройничного нерва. Все ядра латеральной зоны (за исключением ретикулярного латерального ядра продолговатого мозга) состоят из нейронов малой и средней величины и лишены крупных элементов. В этой зоне располагаются восходящие и нисходящие пути, обеспечивающие связь сенсорных образований с медиальной зоной ретикулярной формации и моторными ядрами ствола. Эта часть ретикулярной формации является более молодым и возможно прогрессивнее, с ее развитием связан факт уменьшения объема осевой ретикулярной формации в ходе эволюционного развития. Таким образом, латеральная зона – это совокупность элементарных интегративных единиц, сформированных вблизи и внутри специфических сенсорных систем.
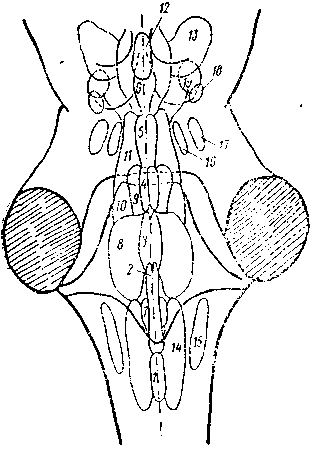
Рис. 2. Ядра ретикулярной формации (РФ) (по: Niuwenhuys еt. аl, 1978).
1—6 — медианная зона РФ: 1—4— ядра шва (1 — бледное, 2 — темное, 3 — большое, 4— мостовое), 5 — верхнее центральное, 6 — дорсальное ядро шва, 7—13 — медиальная зона РФ: 7 — ретикулярное парамедианное, 8 — гигантоклеточное, 9 — ретикулярное ядро покрышки моста, 10, 11 — каудальное (10) и оральное (11) ядра моста, 12 — дорсальное покрышечное ядро (Гуддена), 13 — клиновидное ядро, 14— I5—латеральная зона РФ: 14 — центральное ретикулярное ядро продолговатого мозга, 15 — латеральное ретикулярное ядро, 16, 17 — медиальное (16) и латеральное (17) парабрахиальные ядра, 18, 19 — компактная (18) и рассеянная (19) части педункуло-понтийного ядра.
|
Благодаря нисходящим влияниям ретикулярная формация оказывает тоническое влияние и на мотонейроны спинного мозга, что в свою очередь повышает тонус скелетной мускулатуры, совершенствует систему обратной афферентной связи. В результате любой двигательный акт совершается значительно эффективнее, осуществляет более точный контроль за движением, но чрезмерное возбуждение клеток ретикулярной формации может привести к дрожанию мышц.
В ядрах ретикулярной формации находятся центры сна и бодрствования, и стимуляция тех или иных центров приводит или к наступлению сна, или к пробуждению. На этом основано применение снотворных. В ретикулярной формации расположены нейроны, реагирующие на болевые раздражения, идущие от мышц или внутренних органов. В ней также расположены специальные нейроны, которые обеспечивают быструю реакцию на внезапные, неопределенные сигналы.
Ретикулярная формация тесно связана с корой больших полушарий, благодаря этому формируется функциональная связь между внешними отделами ЦНС и стволом головного мозга. Ретикулярная формация играет важную роль как в интеграции сенсорной информации, так и в контроле над деятельностью всех эффекторных нейронов (моторных и вегетативных). Она имеет также первостепенное значение для активации коры больших полушарий, для поддержания сознания.
Необходимо отметить, что кора полушарий большого мозга, и в свою очередь, посылает по корково-ретикулярным путям импульсы в ретикулярную формацию. Эти импульсы возникают в основном в коре лобной доли и проходят в составе пирамидных путей. Корково-ретикулярные связи оказывают либо тормозное, либо возбуждающее действие на ретикулярную формацию ствола головного мозга, они осуществляют корректировку прохождения импульсов по эфферентным путям (отбор эфферентной информации).
Таким образом, между ретикулярной формацией и корой полушарий большого мозга имеется двусторонняя связь, которая обеспечивает саморегуляцию в деятельности нервной системы. От функционального состояния ретикулярной формации зависит тонус мускулатуры, работа внутренних органов, настроение, концентрация внимания, память и т. д. В целом ретикулярная формация создает и поддерживает условия для осуществления сложной рефлекторной деятельности с участием коры полушарий большого мозга.
Конечный мозг.
Конечный мозг (telencephalon) представлен двумя полушариями (hemispheri cerebri). В состав каждого полушария входят плащ, или мантия (pallium), обонятельный мозг (rhinencephalon) и узлы основания (базальные ганглии). Остатком первоначальных полостей обоих пузырей конечного мозга являются боковые желудочки (ventriculi lateralis). Передний мозг, из которого выделяется конечный, вначале возникает в связи с обонятельным рецептором (обонятельный мозг), а затем становится органом управления поведением животного, причем в нем возникают центры инстинктивного поведения, основанного на видовых реакциях (безусловные рефлексы), - подкорковые узлы, и центры индивидуального поведения, основанного на индивидуальном опыте (условные рефлексы), - кора большого мозга. Соответственно этому в концевом мозге различают в порядке исторического развития следующие группы центров:
Обонятельный мозг - самая древняя и вместе с тем самая меньшая часть, расположенная вентрально.
Базальные, или центральные, ядра полушарий, «подкорка», старая часть конечного мозга (paleencephalon), скрытая в глубине.
Новая кора (cortex) - самая молодая часть (neоencephalon) и вместе с тем самая большая часть, покрывающая остальные как плащом, откуда и ее название - плащ, или мантия.
Так как в процессе эволюции из всех отделов ЦНС быстрее и больше всего растет конечный мозг, у человека он становится самой крупной частью головного мозга и приобретает вид двух объемных полушарий - правого и левого.
В глубине продольной щели мозга оба полушария соединены толстой горизонтальной пластинкой - мозолистым телом (corpus collosum), которое состоит из нервных волокон, идущих поперечно из одного полушария в другое. В мозолистом теле различают загибающийся книзу конец, или колено), среднюю часть, и задний конец, утолщенный в форме валика. Все эти части хорошо видны на продольном разрезе мозга между полушариями. Колено мозолистого тела, загибаясь книзу, заостряется и образует клюв, который переходит в тонкую пластинку, продолжающуюся в свою очередь в конечную пластинку.
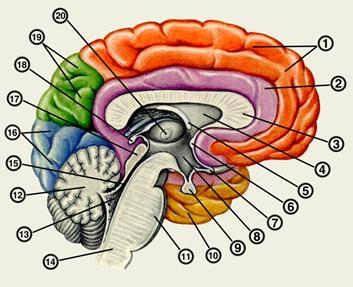
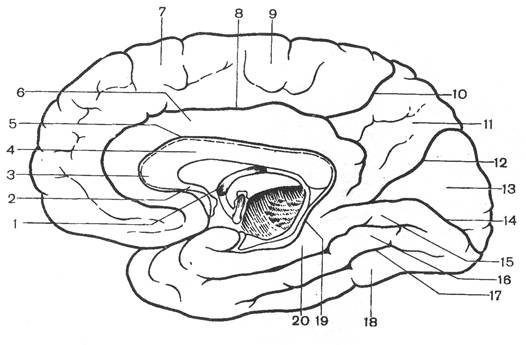
Рис. 1. Сагиттальный разрез головного мозга:
1 - лобная доля; 2 - поясная извилина; 3 - мозолистое тело; 4 - прозрачная перегородка; 5 - свод; 6 - передняя спайка; 7 - зрительный перекрест; 8 - подталамическая область; 9 - гипофиз; 10 - височная доля; 11 - мост; 12 - продолговатый мозг; 13 - четвертый желудочек; 14 - мозжечок; 15 - водопровод мозга; 16 - затылочная доля; 17 - пластинка крыши; 18 - шишковидное тело; 19 - теменная доля; 20 - таламус.
Под мозолистым телом находится так называемый свод (fornix), представляющий два дугообразных белых тяжа, которые в средней своей частисоединены между собой, а спереди и сзади расходятся, образуя впереди столбики свода, позади же - ножки свода. Ножки свода, направляясь назад, спускаются в нижние рога боковых желудочков и переходят в бахромку гиппокампа. Между ножками свода под задним концом колена мозолистого тела протягиваются поперечные пучки нервных волокон, образующие комиссуру свода. Передние концы свода продолжаются в них до основания мозга, где оканчиваются в сосочковых телах, проходя через серое вещество гипоталамуса. Столбики свода ограничивают лежащие позади них межжелудочковые отверстия, соединяющие III желудочек с боковыми желудочками. Впереди столбиков свода находится передняя спайка, имеющая вид белой поперечной перекладины, состоящей из нервных волокон. Между передней частью свода и коленом натянута тонкая вертикальная пластинка мозговой ткани - прозрачная перегородка.
Кора полушарий большого мозга. Кора полушарий большого мозга представляет собой слой серого вещества, толщина которого в различных отделах неодинакова и в среднем равняется 2—3 мм. Поверхность коры имеет сложный рельеф, характеризующийся многочисленными бороздами, и расположенными между ними возвышениями -извилинами. Извилины между собой различаются по форме и величине, однако одноименные извилины на коре полушарий у различных людей принципиально сходны и локализуются в определенных местах. Площадь коры взрослого человека составляет около 220000 мм2, причем 2/3 залегает в глубине между извилинами и только 1/3 лежит на поверхности.
В каждом полушарии большого мозга различают:
- медиальную,
- дорсо-латеральную,
- нижнюю поверхности.
Дорсо-латеральная поверхность полушарии выпукла, наиболее обширна, обращена вверх и латерально, граничит с медиальной поверхностью ясно выраженным краем.
Плоская медиальная поверхность обращена к средней линии, в средней части соединена мозолистым телом с такой же поверхностью другого полушария.
Нижняя поверхность в переднем отделе уплощена, а в заднем вогнута.
Три основные борозды делят каждое полушарие на четыре доли: лобную, теменную, височную, затылочную и островок.
Рассмотрим рельеф коры полушарий большого мозга.
Рис. 2. Верхнелатеральная поверхность полушария:1 – латеральная борозда; 2 – средняя лобная извилина; 3 – верхняя лобная извилина; 4 – прецентральная извилина; 5 – верхняя и нижняя прецентральные борозды; 6 – центральная борозда; 7 – постцентральная извилина; 8 – постцентральная борозда; 9 – внутритеменная борозда; 10 – верхнетеменная долька; 11 – нижнетеменная долька; 12 – надкраевая извилина; 13 – угловая извилина; 14 – затылочный полюс; 15 – нижняя височная борозда; 16 – нижняя височная извилина; 17 – средняя височная извилина; 18 – верхняя височная извилина; 19 – верхняя височная борозда; 20 нижняя лобная борозда; 21 нижняя лобная извилина; 22 – верхняя лобная борозда; 23 – мозжечок; 24 – продолговатый мозг.
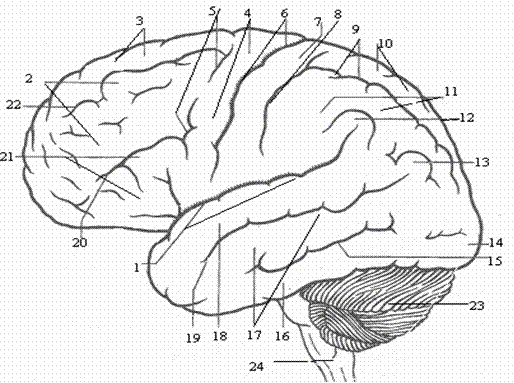
В переднем отделе каждого полушария большого мозга находится лобная доля.
Она заканчивается спереди лобным полюсом и ограничена снизу латеральной бороздой (сильвиева борозда), а сзади - глубокой центральной бороздой. Латеральная борозда, начинаясь на нижней поверхности полушария, идет по латеральной стороне вверх и потом назад, разделяя лобную и височную доли. Центральная борозда расположена во фронтальной плоскости. Она начинается в верхней части медиальной поверхности полушария большого мозга, рассекает его верхний край, спускается, не прерываясь, по верхнелатеральной поверхности полушария вниз и заканчивается, немного не доходя до латеральной борозды. Она отделяет лобную долю от теменной и височной. Спереди от центральной борозды, почти параллельно ей, располагается предцентральная борозда. Последняя заканчивается внизу, не доходя до латеральной борозды. Предцентральная борозда нередко прерывается в средней части и состоит из двух самостоятельных борозд. От предцентральной борозды вперед направляются верхняя и нижняя лобные борозды. Они расположены почти параллельно друг другу и делят верхнелатеральную поверхность лобной доли на извилины. Между центральной бороздой сзади и предцентральной бороздой спереди находится предцентральная извилина. Выше верхнелобной борозды лежит верхняя лобная извилина. Между верхней и нижней лобными бороздами тянется средняя лобная извилина. Книзу от нижней лобной борозды расположена нижняя лобная извилина. В эту извилину снизу вдаются ветви латеральной борозды: восходящая ветвь и передняя ветвь, которые делят нижнюю часть лобной доли, на три части: покрышечную часть (лобная покрышка), прикрывающую лежащую в глубине борозды островковую долю (островок); треугольную часть и глазничную часть.
Кзади от центральной борозды находится теменная доля. Задней границей этой доли является теменно-затылочная борозда. Эта борозда располагается на медиальной поверхности полушария, глубоко рассекает верхний край полушария большого мозга и переходит на его верхнелатеральную поверхность. Границей между теменной и затылочной долями на дорсолатеральной поверхности полушария большого мозга служит условная линия - продолжение теменно-затылочной борозды книзу. Нижней границей теменной доли является латеральная борозда, отделяющая эту долю от височной.
В пределах теменной доли выделяют nocтцентральную борозду. Она начинается от латеральной борозды внизу и заканчивается вверху, не доходя до верхнего края полушария. Постцентральная борозда лежит позади центральной борозды и почти параллельна ей. Между центральной и постцентральной бороздами располагается постцентральна извилина. Вверху она переходит на медиальную поверхность полушария большого мозга, где соединяется с предцентральной извилиной лобной доли, образуя вместе с ней парацентральную дольку. От постцентральной борозды кзади отходит внутритеменная борозда. Она параллельна верхнему краю полушария. Кверху от внутритеменной борозды находится группа мелких извилин, получивших название верхней теменной дольки. Ниже этой борозды лежит нижняя теменная долька, в пределах которой выделяют две извилины: надкраевую, и угловую. Надкраевая извилина охватывает конец латеральной борозды, а угловая — конец верхней височной борозды. Нижняя часть нижней теменной дольки и прилежащие к ней нижние отделы постцентральной извилины вместе с нижней частью предцентральной извилины, нависающие над островковой долей, образуют лобно-теменную покрышку островка.
Затылочная доля, располагается позади теменно-затылочной борозды и ее условного продолжения на верхнелатеральной поверхности полушария. По сравнению с другими долями она имеет небольшие размеры. Затылочная доля кончается затылочным полюсом. Борозды и извилины на верхнелатеральной поверхности затылочной доли очень вариабельны. Наиболее часто и лучше других выражена поперечная затылочная борозда, которая является как бы продолжением кзади внутритеменной борозды теменной доли.
Височная доля, занимает нижнебоковые отделы полушария и отделяется от лобной и теменной долей латеральной бороздой. Край височной доли, прикрывающий островковую долю, получил название височной покрышки. Передняя часть височной доли образует височный полюс. На боковой поверхности височной доли видны две борозды - верхняя и нижняя височные, почти параллельные латеральной борозде. Извилины височной доли ориентированы вдоль борозд. Верхняя височная извилина, расположена между латеральной бороздой вверху и верхней височной внизу. Между верхней и нижней височными бороздами находится средняя височная извилина. Нижнелатеральный край височной доли занимает нижняя височная извилина, ограниченная сверху одноименной бороздой. Задний конец этой извилины продолжается в затылочную долю.
Островковая доля (островок), находится в глубине латеральной борозды. Эту долю можно увидеть, если раздвинуть или удалить прикрывающие островок покрышки лобной, теменной и височной долей. Глубокая круговая борозда островка отделяет островок от окружающих его отделов мозга.
На медиальной поверхности над мозолистым телом, отделяя его от остальных отделов полушария, находится борозда мозолистого тела. Огибая сзади валик мозолистого тела эта борозда направляется книзу и вперед и продолжается в гиппокампальную борозду. Выше борозды мозолистого тела находится поясная борозда. Эта борозда начинается кпереди и книзу от клюва мозолистого тела, поднимается вверх, затем поворачивает назад и следует параллельно борозде мозолистого тела, заканчивается выше и кзади от валика мозолистого тела как подтеменная борозда. Между бороздой мозолистого тела и поясной бороздой находится поясная извилина, охватывающая мозолистое тело спереди, сверху и сзади. Сзади и книзу от валика мозолистого тела поясная извилина суживается, образуя перешеек поясной извилины. Далее книзу и кпереди перешеек переходит в более широкую парагиппокампальную извилину, ограниченную сверху бороздой гиппокампа. Поясная извилина, перешеек и парагиппокампальная извилина известны как сводчатая извилина. В глубине борозды гиппокампа находится зубчатая извилина.
На медиальной поверхности затылочной доли расположены сливающиеся друг с другом под острым углом, открытым кзади, две глубокие борозды: теменно-затылочная борозда, отделяющая теменную долю от затылочной, и шпорная борозда. Последняя начинается на медиальной поверхности затылочного полюса и направляется вперед до перешейка поясной извилины. Участок затылочной доли, лежащий между теменно-затылочной и шпорной бороздами и имеющий форму треугольника, обращенного вершиной к месту слияния этих борозд, называется клином. Хорошо заметная на медиальной поверхности полушария шпорная борозда ограничивает сверху язычную извилину, простирающуюся от затылочного полюса сзади до нижней части перешейка поясной извилины. Снизу от язычной извилины располагается коллатеральная борозда, принадлежащая уже нижней поверхности полушария.
Рис. 3. Медиальная поверхность полушария:1 – свод; 2 - клюв мозолистого тела; 3 - колено мозолистого тела; 4 - ствол мозолистого тела; 5 - борозда мозолистого тела; 6 - поясная извилина; 7 - верхняя лобная извилина; 8 -, 10 - поясная борозда; 9 - парацентральная долька; 11 – предклинье; 12 - теменно-затылочная борозда; 13 – клин; 14 - шпорная борозда; 15 - медиальная затылочно-височная извилина; 16 - средняя затылочно-височная извилина; 17 - затылочно-височная борозда; 18 - боковая затылочно-височная извилина; 19 - гиппокампова борозда; 20 - парагиппокамповая извилина
Рельеф нижней поверхности полушария очень сложен. Передние отделы этой поверхности образованы лобной долей полушария, позади которой выступает височный полюс, а также находятся нижние поверхности височной и затылочной долей, переходящие одна в другую без заметных границ. На нижней поверхности лобной доли, несколько латеральнее и параллельно продольной щели большого мозга, находится обонятельная борозда. Снизу к ней прилежат обонятельная луковица и обонятельный тракт, переходящий сзади в обонятельный треугольник. Участок лобной доли между продольной щелью большого мозга и обонятельной бороздой получил название прямой извилины. Поверхность лобной доли, лежащая латеральнее обонятельной борозды, разделена неглубокими глазничными бороздами, на несколько вариабельных по форме, расположению и размерам глазничных извилин.
Рис. 4. Нижняя поверхность полушария:1- прямая извилина; 2 – обонятельная борозда; 3 – глазничные борозды; 4 – глазничные извилины; 5 - переднее продырявленное вещество; 6 – височно-затылочная борозда; 7 латеральная височно-затылочная извилина; 8 – медиальная височно-затылочная извилина; 9 – коллатеральная борозда; 10 - гиппокампова борозда; 11 - медиальная затылочно-височная извилина; 12 - шпорная борозда; 13 - парагиппокамповая извилина; 14 – крючок; 15 – сосцевидные тела; 16 – средний мозг; 17 – обонятельная луковица; 18 – обонятельный тракт; 19 – зрительный перекрест
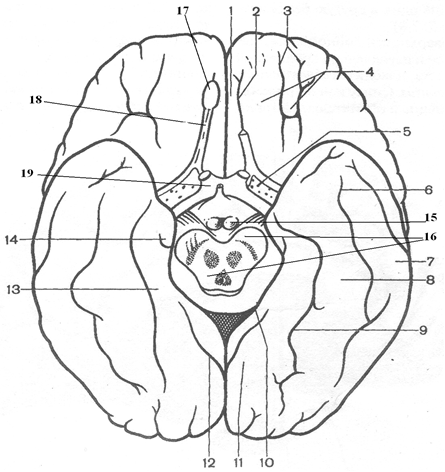
В заднем отделе нижней поверхности полушария хорошо различима коллатеральная борозда, лежащая книзу и латерально от язычной извилины на нижней поверхности затылочной и височной долей, латерально от парагиппокампальной извилины, Несколько кпереди от переднего конца коллатеральной борозды находится носовая борозда, sulcus rhindlis. Она ограничивает с латеральной стороны изогнутый конец парагиппокампальной извилины - крючок. Латеральнее коллатеральной борозды лежит медиальная затылочно-височная извилина. Между этой извилиной и расположенной кнаружи от нее латеральной затылочно-височной извилиной, находится затылочно-височная борозда.
Обонятельный мозг. Обонятельный мозг (rhinencephalon) — филогенетически самая древняя часть переднего мозга, возникшая в связи с рецептором обоняния, когда передний мозг не стал еще органом поведения животного. Поэтому все компоненты его являются различными частями обонятельного анализаора. Обонятельный мозг располагается на нижней и медиальной поверхностях полушарий мозга и условно разделяется на периферический и центральный отделы.
К периферическому отделу обонятельного мозга относятся обонятельная луковица и обонятельный тракт, находящиеся на нижней поверхности лобной доли в обонятельной борозде. Обонятельный тракт заканчивается обонятельным треугольником, который впереди переднего продырявленного вещества расходится двумя обонятельными полосками. Латеральная полоска заканчивается в коре крючка височной доли. Медиальная полоска направляется в подмозолистую извилину и околообонятельное поле, которые располагаются под клювом мозолистого тела.
К центральному отделу обонятельного мозга относятся: сводчатая извилина, гиппокамп, зубчатая извилина и крючок.
Гиппокамп - парное образование, представляет впячивание серого вещества со стороны медиальной стенки нижнего рога бокового желудочка. Гиппокамп хорошо виден в полости нижнего рога в виде булавовидного тела. В гиппокамп диффузно проецируются многие афферентные системы, эфферентные же влияния направлены преимущественно к гипоталамусу. Полагают, что гиппокамп играет существенную роль в поддержании постоянства внутренней среды организма, участвует в высшей координации функций размножения и эмоционального поведения, а также в процессах обучения и сохранения памяти. Гиппокамп является также центром обоняния.
Базальные ганглии располагаются в глубине белого вещества полушарий. К ним относят полосатое тело, состоящее хвостатого и чечевицеобразного ядер, миндалевидное ядро и ограду. Эти ядра разобщены друг от друга прослойками белого вещества, образующими внутреннюю, наружную и крайнюю капсулы.
Рис. 5. Базальные (подкорковые) ядра на фронтальном разрезе головного мозга: 1-сосудистое сплетение бокового желудочка (центральная часть); 2-таламус; 3-внутренняя капсула; 4-кора островка; 5-ограда; 6-миндалевидное тело; 7-зрительный тракт; 8-сосцевидное тело; 9-бледный шар; 10-скорлупа; 11-свод мозга; 12-хвостатое ядро; 13-мозолистое тело.
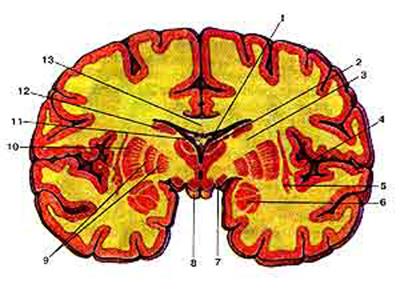
Полосатое тело разделяется пучком нервных волокон, идущих от коры и называемых внутренней капсулой, на две части хвостатое ядро и скорлупу. Хвостатое ядро имеет булавовидную форму и изогнуто назад. Передняя его часть расширена, называется головкой и располагается выше чечевицеобразного ядра, а его задняя часть - хвост проходит сверху и латеральнее таламуса, отделяясь от него мозговыми полосками. Головка хвостатого ядра участвует в образовании латеральной стенки переднего рога бокового желудочка. Хвостатое ядро состоит из малых и больших пирамидных клеток.
Чечевицеобразное ядро находится латерально и кпереди от таламуса и хвостатого ядра. На фронтальном срезе имеет форму треугольника. Две параллельные вертикальные прослойки белого вещества делят чечевицеобразное ядро на 3 части: скорлупу (самая латеральная часть) и медиальную и латеральную пластинки бледного шара. Хвостатое ядро и скорлупа являются филогенетически новыми образованиями, их объединяют под общим названием неостриатум. Бледный шар - более древнее образование, носит название палеостриатум или паллидум. Вместе они образуют так называемую стриопаллидарную систему.
Полосатое тело получает афферентные импульсы главным образом от таламуса, отчасти от коры; посылает эфферентные импульсы к бледному шару. Полосатое тело рассматривают как эффекторное ядро, не имеющее самостоятельных двигательных функций, но контролирующее функции филогенетически более старого двигательного центра - бледного шара. Полосатое тело регулирует и частично затормаживает безусловнорефлекторную деятельность бледного шара, т. е. действует на него так же, как бледный шар действует на красное ядро. Полосатое тело считают высшим подкорковым регуляторно-координационным центром двигательного аппарата. В полосатом теле, согласно экспериментальным данным, находятся также высшие вегетативные координационные центры, регулирующие обмен веществ, теплообразование и тепловыведение, а также сосудистые реакции. По-видимому, в полосатом теле находятся центры, которые интегрируют, объединяют безусловнорефлекторные двигательные и вегетативные реакции в единый целостный акт поведения.
При поражениях полосатого тела у человека наблюдается атетоз — стереотипические движения конечностей, а также хорея — сильные неправильные движения, совершающиеся без всякого порядка и последовательности и захватывающие почти всю мускулатуру («пляска святого Витта»). И атетоз и хорея рассматриваются как результат выпадения тормозящего влияния, которое оказывает полосатое тело на бледное ядро.
Бледный шар - парное образование, входящее в состав чечевицеобразного ядра, и являющееся двигательным ядром. При его раздражении можно получить сокращение шейных мышц, конечностей и всего туловища, преимущественно на противоположной стороне. Бледное ядро получает импульсы по афферентным волокнам, идущим от таламуса и замыкающим таламопаллидарную рефлекторную дугу. Бледное ядро, будучи связано эффекторно с центрами среднего и заднего мозга, регулирует и координирует их работу. Одной из функций бледного ядра считают торможение нижележащих ядер, главным образом красного ядра среднего мозга, в связи с чем при повреждении бледного шара наблюдается сильное увеличение тонуса скелетной мускулатуры — гипертонус, так как красное ядро освобождается от тормозящего влияния бледного шара. Таламо-гипоталамо-паллидарная система принимает участие у высших животных и человека в осуществлении сложных безусловных рефлексов — оборонительных, ориентировочных, пищевых, половых.
Миндалевидное ядро представляет группу ядер и локализуется внутри переднего полюса височной доли, латеральнее перегородки продырявленного вещества. В функциональном отношении является частью лимбической системы, участвует в регуляции вегетативных и нейроэндокринных реакций. Миндалина характеризуется очень низким порогом возбуждения, что может способствовать развитию эпилептиформной активности. При стимуляции миндалины возникают судороги, эмоционально окрашенные реакции, страх, агрессия и т. д.
Ограда - тонкая прослойка серого вещества, отделенная наружной капсулой белого вещества от чечевицеобразного ядра. Ограда внизу соприкасается с ядрами переднего продырявленного вещества. Предполагают участие в осуществлении глазодвигательных реакций слежения за объектом.
Между хвостатым ядром и таламусом с одной стороны и чечевицеобразным ядром - с другой находится прослойка белого вещества, называемая внутренней капсулой. В ней проходят все проекционные волокна к коре полушарий большого мозга и от коры к нижележащим отделам ЦНС. Ее подразделяют на 3 отдела: переднюю ножку, колено и заднюю ножку.
В передней ножке внутренней капсулы проходят волокна, образованные нейронами лобных областей коры: лобно-таламические (tr. frontothalamicus), лобно- красноядерные (tr. frontorubralis) и лобно-мостовые(tr. frontopontinus) пути.
В колене внутренней капсулы располагается корково-ядерный путь (tr. corticonuclearis).
Заднюю ножку в переднезаднем направлении образуют: корково-спинномозговой (tr. corticospinalis), таламо-кортикальный (tr. thalamocorticalis), затылочно-височно-мостовой (tr. occipitotemporopontinus), слуховая лучистость (radiatio acustica), зрительная лучистость (radiatio optica). Вниз волокна направляются в ножки среднего мозга. Выше внутренней капсулы волокна образуют лучистый венец.
Внутреннее строение новой коры. У человека в коре выделяют шесть слоев:
1 - молекулярная пластинка,
2 - наружная зернистая пластинка,
3 - наружная пирамидная пластинка,
4 - внутренняя зернистая пластинка.
5 - внутренняя пирамидная пластинка,
6 - мультиформная пластинка.
Рис. 6. Строение новой коры. I - молекулярная пластинка, II - наружная зернистая пластинка, III - наружная пирамидная пластинка, IV - внутренняя зернистая пластинка, V - внутренняя пирамидная пластинка, VI - мультиформная пластинка.

Молекулярная пластинка, является самым наружным слоем коры, бедна клеточными элементами. Здесь расположена густая сеть, образованная дендритами пирамидных нейронов и аксонов клеток других слоев. Главное назначение этого слоя - обеспечение межнейронных связей клеток различных слоев.
Наружная зернистая пластинка состоит из звездчатых нейронов и малых пирамид. В этом слое происходит дихотомическое ветвление дендритов пирамидных нейронов, и проходит много горизонтальных волокон. Основная функция - формирование вертикальных связей.
Наружный пирамидный слой содержит пирамидные клетки разного размера. Аксоны их не образуют длинных проводящих путей. На нейронах этого слоя заканчиваются ассоциативные афференты.
Внутренняя зернистая пластинка состоит из плотно расположенных звездчатых нейронов. Здесь заканчиваются таламокортикальные волокна.
Внутренняя пирамидная пластинка содержит крупные и гигантские пирамиды. Их апикальные дендриты поднимаются в первый слой. От этого слоя начинаются корково-ядерные и корково-спинальные пути.
Мультиформная пластинка содержит нейроны переходных форм разных размеров и продолжается в белое вещество без резкой границы. Обеспечивает восходящие и горизонтальные связи.
Функциональной единицей коры является вертикальная колонка, состоящая из 3-7 клеток, они вместе реагируют на один и тот же раздражитель.
Локализация функций в новой коре. Вид, взаимное расположение нейронов неодинаковы в различных областях коры. Цитоархитектонические исследования (исследования расположения нейронов) позволили произвести картирование коры. Общепринятой является классификация полей К. Бродмана (1909 г.), которая предусматривает деление коры на 52 поля и цифровое обозначение последних. Эта нумерация легла в основу цитоархитектонической карты, составленной Институтом мозга РАН. В ней рад полей подразделяется на зоны, обозначаемые латинскими буквами.
Рис. 7. Цитоархитектоническая карта коры больших полушарий.
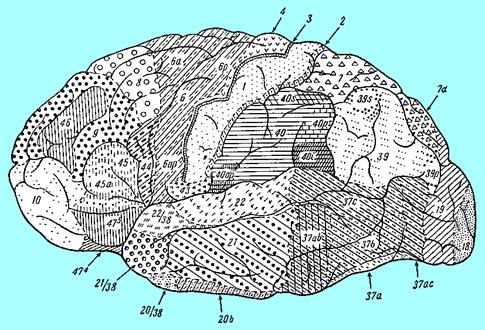
В настоящее время установлено функциональное значение различных областей коры. Участки коры с определенной цитоархитектоникой и характерными нервными связями, участвующие в выполнении определенных функций, называются нервным центрами. Традиционно центры новой коры принято делить на проекционные (первичные и вторичные) и ассоциативные. Проекционные центры - участки коры, являющиеся корковой частью того или иного анализатора. Критерий отнесения центров к первичным - существование прямого входа от проекционных ядер таламуса. Для них характерны строгая топологическая организация входов и пропорциональная зависимость площади представительства от плотности иннервации соответствующего участка рецептурной поверхности. Следствие повреждения первичной проекционной зоны - выпадение восприятия стимулов, приходящих на соответствующий участок рецепторной поверхности.
Вторичные зоны располагаются вблизи первичных проекцонных центров и являются их периферическими отделами. Для них характерно помимо прямых входов от проекционных таламических ядер, входы от первичных проекционных центров, а также преимущественное представительство наиболее интенсивно иннервируемых, а следовательно наиболее важных в функциональном отношении отделов. Роль вторичных полей в процессах восприятия и организации движений оказывается более сложной по сравнению с первичными. Повреждения приводят к нарушению сложных форм восприятия, распознавания и оценки стимулов.
Ассоциативные центры в мозге человека занимают более половины всей поверхности полушарий большого мела и являются наиболее молодыми образованиями. Ассоциативные центры связаны с ассоциативными ядрами таламуса и с проекционными центрами коры. Ассоциативные центры принимают участие в организации сложных форм поведения, в осуществлении высшей нервной деятельности. Анатомически и функционально ассоциативные центры часто асимметричны.
Основными проекционными центрами являются:
1. Центр общей чувствительности (тактильной, температурной, болевой, осознаваемой пропроицептивной). Локализуется в постцентральной извилине (поля 3 - первичная зона; 1,2 - вторичная зона). Поля соматотопически организованы. В верхнем отделе постцентральной извилины проецируются туловище и нижняя конечность, в среднем - верхняя конечность, в нижнем - голова. Поражение центра сопровождается утратой тактильной, температурной, болевой чувствительности и мышечно-суставного чувства на противоположной половине тела.
2. Центр двигательных функций занимает поле 4 предцентральной извилины (первичная зона) и поле 6 парацентральной дольки (вторичная зона). Здесь осуществляется анализ проприоцептивных раздражений. От нейронов внутреннего пирамидного слоя берут начало пирамидные тракты. В поле 4 имеется четкая соматотопическая организация - "моторный гомункулус Пенфилда". Тело проецируется "вверх ногами" на кору противоположного полушария. Поражение зоны ведет к нарушению восприятия проприоцептивных раздражений, могут возникнуть центральные параличи. Центр двигательных функций необходим для выполнения интегративных функций при выполнении произвольных движений.
3. Центр схемы тела расположен в теменной доле (поле 40). В нем представлены соматотопические проекции всех частей тела. Сюда поступает осознаваемая проприоцептивная чувствительность. Назначение центра - определение положения тела и его частей в пространстве и оценка тонуса мускулатуры. Нарушение центра ведет к невозможности узнавания частей собственного тела, ощущению лишних конечностей, нарушению определения положения тела и его частей в пространстве.
4. Центр зрения расположен в затылочной доле (поле 17 - первичная зона, поля18, 19 - вторичная). На нейронах 17 поля спроецирована сетчатка. Нейроны 18 поля обеспечивают зрительную память, а 19 - ориентацию в непривычной обстановке. Одностороннее поражение поля 17 сопровождается частичной слепотой на оба глаза, но в различных участках сетчатки. Поражение 18 и 19 полей приводит к искаженному зрительному восприятию.
5. Центр слуха расположен в верхней височной извилине, на поверхности, обращенной к островку (поле 41). Это первичный слуховой центр, одностороннее поражение которого приводит к снижению слуха на оба уха, на противоположной стороне - в большей степени. Двустороннее повреждение приводит к полной глухоте.
6. Центр вкуса располагается на медиальной поверхности височной доли (поля 11, А, Е). Здесь заканчиваются волокна вкусового пути своей и противоположной сторон. Эти области относят к лимбической доле мозга, поражение которой вызывает расстройство вкуса, обоняния, появлению галлюцинаций.
7. Центр обоняния располагается там же, где и проекционный центр вкуса. Здесь заканчиваются волокна обонятельного пути своей и противоположной сторон. Одностороннее повреждение приводит к снижению обоняния и обонятельным галлюцинациям.
8. Центр вестибулярных функций расположен на дорсальной поверхности височной доли (поля 20,21,22). Поражение указанных отделов приводит к спонтанному головокружению, ощущению неустойчивости, чувству проваливания, ощущению деформации окружающих предметов и их движения.
9. Центр висцерорецепции занимает поле 43 нижней трети постцентральной и предцентральной извилин. Сюда поступает информация от интерорецепторов внутренних органов. В центре анализируются главным образом болевые ощущения.
Основными ассоциативными центрами являются:
1. Центр стереогнозии (узнавания предметов на ощупь). Располагается в верхней теменной дольке (поле 7). Функция центра - узнавание ранее встречавшихся предметов. Центр постоянно развивается. При поражении центра теряется способность с закрытым [глазами создать общее целостное представление о предмете, тогда как отдельные свойства (форма, фактура, масса, температура и т.п.) определяются правильно.
2. Центр праксии (целенаправленных привычных движений). Располагается в нижней теменной дольке (поле 40) у правшей - в левом полушарии, у левшей - в правом. Амбидексы (одинаково владеющие обеими руками) имеют центр в обоих полушариях. Центр развивается в результате многократного повторения сложных целенаправленных движений. Поражение приводит к утрате произвольных приобретенных практикой движений.
3. Центр зрительной памяти. Располагается на дорсальной поверхности затылочной доли (поля 18-19) у правшей - слева, у левшей – справа. Обеспечивает запоминание предметов по их форме, внешнему виду, цвету. Поражение центра ведет к зрительной агнозии. Может наблюдаться частичная агнозия (не узнает знакомых, свой дом, себя на фото).
Центры, связанные с речевой функцией.
4. Акустический центр речи (центр Вернике). Расположен в области верхней височной извилины (поле 42). Поражение центра приводит к сенсорной афазии (словесная глухота). Хотя больной слышит, он не понимает речь. Нарушается слуховой контроль собственной речи, что ведет к невозможности построения связных предложений. Речь таких больных представляет набор бессмысленных слов и звуков.
5. Центр моторной речи (центр Брока). Располагается в области нижней лобной извилины (поле 44) у правшей – слева, у левшей – справа. При поражении развивается моторная афазия – неспособность говорить при полном сохранении понимания и внутренней речи.
6. Центр речевого анализатора пения. Расположен рядом с предыдущим (центральные отделы нижней лобной извилины) (поле 45). Поражение центра сопровождается вокальной амузией – неспособность к восприятию и составлению музыкальных фраз, и аграмматизмом – неспособность составления осмысленных предложений из отдельных слов. Речь больных – несвязанный набор слов.
7. Зрительный анализатор письменной речи. Расположен в угловой извилине нижней теменной дольки (поле 39). В центре проходит анализ зрительной информации о буквах, цифрах, составе слов и понимание их смысла. Поражение ведет к неспособности читать – алексии. Больной видит буквы, но не понимает смысла.
8. Двигательный анализатор письменной речи. Занимает задние отделы среднелобной извилины (поле 8). При поражении центра возникает аграфия (неспособность производить рукой точные и тонкие движения, необходимые при письме).
Эти центры развиваются только у человека и совершенствуются всю жизнь.
Слуховой и двигательный центры речи закладываются на 3-4 месяце жизни. Зрительный центр речи – на четвертом году жизни. Двигательный центр письма начинает формироваться на 5-6 году жизни.
Кора головного мозга, подкорковые структуры, а также периферические компоненты организма связаны волокнами нейронов, образующими несколько типов проводящих путей. Ассоциативные волокна — проходят внутри только одного полушария и связывают соседние извилины в виде коротких дугообразных пучков, либо кору различных долей, что требует более длинных волокон. Назначение ассоциативных связей — обеспечение целостной работы одного полушария как анализатора и синтезатора разномодальных возбуждений. Проекционные волокна — связывают периферические структуры с корой головного мозга. Восходящие проводящие пути. — передют информацию к соответствующим корковым представительствам того или иного анализатора. Нисходящие волокна — начинаются от двигательных зон мозга. Задача этих волокон — организация двигательной активности. Комиссуральные волокна — обеспечивают целостную совместную работу двух полушарий. Они представлены одним крупным анатомическим образованием — мозолистым телом, а также несколькими более мелкими структурами, важнейшими из которых являются свод, передняя и задняя комиссура.
Дата добавления: 2015-11-28; просмотров: 4303;