Белое вещество мозжечка
Включает белое вещество, лежащее в толще мозжечка, и три пары ножек. В толще мозжечка расположены волокна, идущие от коры мозжечка к его ядрам, а также продолжение афферентных волокон, образующих ножки мозжечка и эфферентные волокна, переходящие в его ножки.
Как уже было сказано, три пары ножек соединяют мозжечок с другими структурами ЦНС.
Нижние ножки связывают мозжечок с продолговатым и спинным мозгом. В них проходят главным образом афферент ные волокна:
1) оливо-мозжечковый путь;
2) задний (дорсальный) спинно-мозжечковый путь;
3) вестибуломозжечковый путь (от вестибулярных ядер мозгового ствола);
4) волокна от нежного и клиновидного ядер продолговатого мозга;
5) волокна от ретикулярной формации.
Проходят в нижних ножках и эфферентные волокна — они начинаются в ядрах шатра и идут к вестибулярным ядрам. От вестибулярных ядер, в свою очередь, начинается вестибулоспинальный тракт.
Средние ножки связывают ядра моста с корой мозжечка (мосто-мозжечковый тракт). Напомним, что ядра моста в свою очередь получают афференты от коры больших полушарий (см. 7.2.3). Таким образом, через средние ножки мозжечок получает информацию о запускаемых большими полушариями двигательных программах.
Верхние ножки содержат главным образом эфферентные волокна, идущие от ядер мозжечка (кроме ядер шатра) к таламусу, красному ядру, РФ. Афферентные волокна верхних ножек — это передний (вентральный) спинно-мозжечковый путь.
Передний мозг
Промежуточный мозг
Передний мозг состоит из двух частей — промежуточного мозга и конечного мозга.
Промежуточный мозг, diencephalon, — конечный отдел мозгового ствола, сверху он покрыт большими полушариями, сзади соединен со средним мозгом. Полостью промежуточного мозга является IIIмозговой желудочек. Он расположен по средней линии и на фронтальном (т.е. параллельном лобной поверхности) разрезе имеет вид узкой вертикальной щели.
Промежуточный мозг (рис. 32, 33) состоит из парных образований — таламусов, thalamus (зрительных бугров), примыкающих к ним сверху непарного эпиталамуса (надбугорья, примыкает к таламусу сверху), непарного гипоталамуса (подбугорья, примыкает к таламусу снизу). Кроме того, в промежуточный мозг входит субталамус, который не виден на поверхности мозга и находится в глубине мозгового вещества между гипоталамусом и средним мозгом.
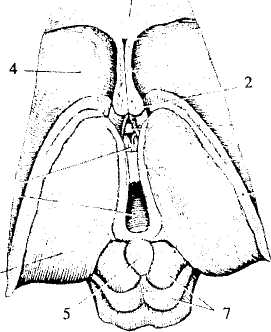
Рис. 32. Промежуточный мозг и средний мозг (вид сверху):
1 — мозолистое тело (конечный мозг); 2 — проводящие пути переднего
мозга; 3 — таламус; 4 — хвостатое ядро (конечный мозг); 5 — эпифиз;
6— III мозговой желудочек; 7— четверохолмие
Таламус.Каждый таламус (см. рис. 32) представляет собой яйцевидное образование длиной примерно 4 см. Медиальные поверхности таламусов образуют боковые стенки третьего желудочка. Между этими стенками находится межбугорное сращение (серое вещество), соединяющее правый и левый таламусы. Передний конец таламуса несколько заострен, а задний расширен и утолщен.
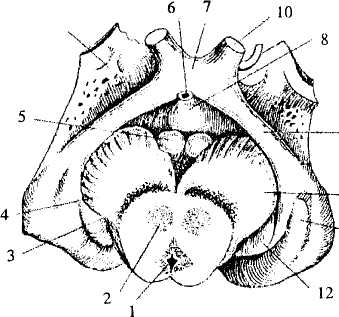
Рис. 33. Промежуточный мозг и средний мозг (вид снизу):
1-4 — средний мозг: 1 — водопровод, 2 — красное ядро, 3 — черная
субстанция; 4 — ножки мозга; 5-8 — гипоталамус: 5 — мамиллярные
тела, 6— воронка, 7— зрительный перекрест, 8— серый бугор;
9— зрительный тракт; 10— зрительный нерв; 11 — 12 — таламус:
11 — наружное коленчатое тело, 12 — внутреннее коленчатое тело;
13— обонятельный треугольник (конечный мозг)
В каждом таламусе расположено около 40 ядер (рис. 34), которые можно разделить по выполняемым ими функциям на проекционные, ассоциативные и неспецифические.
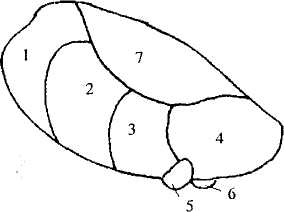
Рис. 34. Ядра таламуса:
1 —лимбическиеядра; 2 — -вентролатеральные (двигательные) ядра;
3— заднее вентральное ядро; 4 — подушка; 5 — латеральное
коленчатое тело; 6— медиальное коленчатое тело;
7—медиодорсалъное (ассоциативное) ядро
Проекционные ядра - это переключательные ядра, полу чающие входы из различных внеталамических структур. Волокна из этих структур образуют синапсы на нейронах проекционных ядер, а аксоны последних проводят импульсацию в определенные локальные области коры больших полушарий. Проекционные ядра подразделяются на сенсорные, двигательные и лимбические.
Сенсорные ядра обеспечивают быстрое проведение сенсорной афферентации от конкретных сенсорных систем в первичные проекционные зоны коры больших полушарий. Пути от всех рецепторов (за исключением обонятельных) проходят через таламус и имеют там свои представительства. Например, в латеральном (наружном) коленчатом теле (ЛКТ), являющемся проекционным зрительным ядром и находящемся в задней наружной части таламуса, заканчиваются волокна зрительного тракта. Из ЛКТ нервные импульсы поступают в затылочную долю коры больших полушарий, где находится центральный отдел зрительного анализатора. В медиальном (внутреннем) коленчатом теле (МКТ) — проекционном слуховом ядре, расположенном в задней внутренней части таламуса, образуют синапсы волокна от слуховых ядер. МКТ посылает свои проекции в слуховую зону коры в височной доле. Отметим, что ЛКТ и МКТ объединяют под названием метаталамус. Проекционным ядром систем кожной и мышечной чувствительности является заднее вентральное ядро таламуса. Здесь заканчиваются волокна от нежного и клиновидного ядер продолговатого мозга (медиальный лемниск) и ядер тройничного нерва. Аксоны клеток заднего вентрального ядра направляются в переднюю часть теменной доли больших полушарий.
Зрительные функции выполняет также одно из ассоциативных ядер таламуса — подушка.
Необходимо подчеркнуть, что в сенсорных ядрах, как и в других ядрах таламуса, происходит не только переключение информации, но и ее обработка. Суть этой обработки состоит в избирательном проведении информации в кору больших полушарий. Иными словами, таламус исполняет роль фильтра, пропуская в конечный мозг либо очень значимые (сильные, новые) сигналы, либо сигналы, связанные с текущей деятельностью коры больших полушарий. Таким образом, таламус является одной из ключевых структур, обеспечивающих и поддерживающих процессы внимания.
Двигательные (моторные) ядра таламуса, лежащие в его нижней боковой части (вентролатеральные ядра), связаны проекционными волокнами с двигательной корой. Они полу чают информацию от мозжечка и базальных ядер, т.е. являются важнейшим переключательным звеном в системе управления движениями.
Лимбические ядра находятся в передней части таламуса. Они входят в лимбическую систему (см. гл. 9) и проводят сен сорную информацию в лимбические отделы коры больших полушарий.
На ассоциативных ядрах таламуса (дорсальная область) оканчиваются волокна не от одной, а сразу от нескольких сенсорных систем, а также от других ядер таламуса и коры больших полушарий. Это обеспечивает их участие в интегративных функциях головного мозга, т.е. в объединении разных видов чувствительности. Эти ядра посылают свои волокна в ассоциативные зоны коры больших полушарий. Дорсальные ядра — эволюционно молодые отделы промежуточного мозга. Их формирование идет параллельно развитию высших (ассоциативных) центров коры.
Неспецифические (медиальные) ядра таламуса, расположенные в его внутренней части, принадлежат, главным образом, ретикулярной системе. Они получают афференты от большого числа образований и посылают диффузные проекции на обширные области коры, влияя, таким образом, на уровень ее активации.
К медиальным ядрам примыкают области таламуса, обеспечивающие обработку и проведение вестибулярной, вкусовой и болевой чувствительности.
Гипоталамус — подбугорная область промежуточного мозга, высший центр регуляции вегетативных и эндокринных функций (см. рис. 20, 21, 33). Он объединяет ряд структур, окружающих нижнюю часть III мозгового желудочка— мамиллярные (сосцевидные) тела, серый бугор, зрительную хиазму. Серый бугор — это непарный полый выступ нижней стенки III желудочка. Его верхушка вытянута в полую воронку, infundibulum, на слепом конце которой находится железа внутренней секреции гипофиз.
С гипоталамусом связан зрительный нерв, который выходя из глазного яблока, входит в полость черепа. В гипоталамической области примерно половина его волокон переходит на другую сторону, образуя зрительную хиазму (перекрест), chiasma opticum. После перекреста зрительные волокна образуют зрительный тракт, волокна которого идут к различным структурам мозга, в частности, к латеральным коленчатым телам таламуса и к некоторым областям гипоталамуса.
Так же, как и в таламусе, в гипоталамусе выделяют несколько десятков ядер. Однако их функциональная классификация пока разработана недостаточно, так как большинство ядер не обладает узкой функциональной специализацией. Топографически выделяют переднюю группу ядер (паравентрикулярное, супраоптическое, супрахиазменное и др.), среднюю группу (ядро воронки или инфундибулярное ядро и др.) и заднюю группу (ядра мамиллярных тел и др.).
Дополнительно к этому в медиолатеральном направлении в гипоталамусе выделяют перивентрикулярную, медиальную и латеральную зоны (рис. 35). Перивентрикулярная зона образована мелкими нейронами, находящимися вдоль стенок III желудочка (греч. peri — вокруг, лат. ventriculus— желудочек). В медиальной зоне находится основная масса гипоталамических ядер. Латеральная зона содержит главным образом белое вещество (волокна, соединяющие гипоталамус с другими структурами ЦНС). Медиальная зона отделена от латеральной важнейшим проводящим пучком переднего мозга — сводом (см. далее).
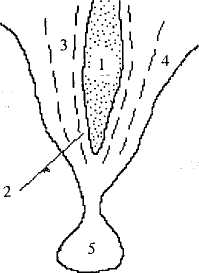
Рис. 35. Зоны гипоталамуса в медиолатеральном направлении:
I — III желудочек; 2 — Перивентрикулярная зона, 3 — медиальная
зона; 4 — латеральная зона; 5— гипофиз
Количество источников афферентации гипоталамуса очень велико. Все его ядра получают прямые входы от коры больших полушарий (особенно ее лобной доли), т.е. аксоны нейронов коры образуют синапсы на клетках ядер гипоталамуса. Также гипоталамус имеет множество сенсорных входов: волокна зрительных нервов, волокна ядер одиночного пути (информация от вкусовых и внутренних рецепторов), волокна латеральной петли (слуховая чувствительность) заканчиваются на его нейронах. Получает афференты гипоталамус от лимбических и ассоциативных ядер таламуса, от РФ, от ряда других образований ЦНС.
Эфференты гипоталамуса главным образом идут к различным исполнительным структурам мозга — вегетативным ядрам, железам внутренней секреции (гипофизу и эпифизу), к покрышке среднего мозга, РФ продолговатого мозга и моста. Некоторые ядра гипоталамуса посылают свои волокна в конечный мозг — к коре больших полушарий и базальным ганглиям.
Большинство этих волокон проходит в составе проводящих пучков переднего мозга.
Отметим только три из них:
1) свод, fornix, волокна, идущие от корковой структуры гиппокампа (см. 7.4.2) к мамиллярным телам гипоталамуса;
2) мамилло-таламический тракт — волокна, идущие от мамиллярных тел к лимбическим ядрам таламуса;
3) гипоталамо-гипофизарный тракт — волокна, идущие от паравентрикулярного и супраоптического ядер к задней доле гипофиза.
Гипоталамус управляет всеми основными гомеостатическими процессами, причем осуществляет это как нервным, так и гуморальным путем.
Нервная регуляция обеспечивается, во-первых, за счет управления деятельностью вегетативной нервной системы и, во-вторых, участием в организации поведения, удовлетворяющего основные биологические потребности организма. Эти функции гипоталамуса обеспечиваются благодаря наличию в нем центров различных потребностей, а также нейронов, реагирующих на изменение внутренней среды организма (температуру крови, ее водно-солевой состав, концентрацию в ней гормонов и т.п.).
Например, при понижении в крови концентрации глюкозы, возбуждается находящийся в сером бугре центр голода, что приводит к возникновению чувства голода. Это вызывает за пуск поведенческих реакций, направленных на удовлетворение пищевой потребности. И наоборот, при повышении в крови концентрации глюкозы (что происходит после еды) возбуждается центр насыщения, тормозящий центр голода. При повышении температуры тела возбуждаются нейроны центра терморегуляции, которые запускают вегетативные ре акции (например, расширение поверхностных кожных сосудов), приводящие к понижению температуры. Также в гипоталамусе находятся центры жажды, водного насыщения, центры полового и родительского поведения (передняя область), цен тры агрессии и страха (задняя область) и т.п. Таким образом, именно здесь определяется уровень актуальности биологических потребностей организма.
Гипоталамус — одна из центральных структур лимбической системы мозга, осуществляющей организацию эмоционально го поведения. Несколько упрощая возникающие при этом процессы, данную функцию гипоталамуса можно описать следующим образом. Если потребности организма удовлетворяются, возбуждается расположенный здесь центр положительного подкрепления, что сопровождается возникновением положи тельных эмоций; если нет— возбуждается центр отрицательного подкрепления, и возникают отрицательные эмоции. Работа систем положительного и отрицательного подкрепления, в свою очередь, лежит в основе процессов обучения в ЦНС, формирования либо ослабления нервных связей (условных рефлексов, ассоциаций).
Гуморальная регуляция осуществляется в тесной связи с гипофизом (см. 1.3.1). Рассмотрим связь между гипоталамусом и гипофизом более подробно.
В нервной ткани есть нейросекреторные клетки, синтезирующие и выделяющие биологически активные вещества, действующие как гормоны. Кроме того, многие медиаторы нервной системы также могут действовать как гормоны. То есть если синтезируемое нейроном вещество выделяется в синоптическую щель и действует на постсинаптическую мембрану, оно является медиатором; если то же вещество выделяется в кровь и действует на орган-мишень — это гормон. Таким образом, нейрогормоны — биологически активные вещества, вырабатываемые нейросекреторными клетками и выделяющиеся в кровь.
Большинство нейрогормонов синтезируется в гипоталамусе — месте непосредственного взаимодействия нервной и эндокринной систем и высшем органе гормональной регуляции основных эндокринных функций. Гипоталамус и гипофиз образуют единую гипоталамо-гипофизарную систему (рис. 36).
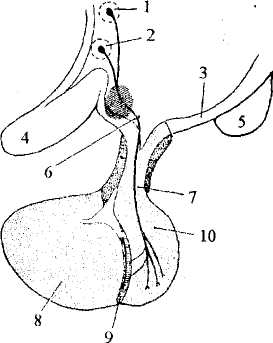
Рис. 36. Гипоталамо-гипофизарная система:
1. — паравентрикулярное ядро; 2 — супраоптическое ядро; 3 — гипофизотропная зона; 4 — зрительный перекрест; 5 — мамиллярные тела;
б— аксоны от нейронов паравентрикулярного и супраоптического ядер,
идущие к гипофизу; 7— воронка; 8-10 — гипофиз: 8 — передняя доля,
9— промежуточная доля, 10— задняя доля
Как уже говорилось, в гипоталамусе есть рецепторы со стояния внутренней среды. Анализируя поступающую ин формацию, гипоталамус трансформирует ее в гуморальные факторы — нейрогормоны. Таким образом, нейроны гипоталамуса активируют либо тормозят выделение гипофизом его гормонов. Рассмотрим этот процесс более детально.
В средней части гипоталамуса (гипофизотропные зоны) находятся мелкоклеточные ядра (ядро воронки, ядра серого бугра), в которых синтезируется пептидные (состоящие из аминокислот) гормоны. Эти гормоны контролируют работу железистых клеток аденогипофиза и промежуточной доли гипофиза. Аксоны нейронов этих ядер оканчиваются в районе воронки, соединяющей гипоталамус и гипофиз. Там они выделяются в кровь и через нее попадают к железистым клеткам передней доли гипофиза. Гормоны, стимулирующие синтез и выделение гормонов гипофиза, получили название рилизинг гормонов (releasing-factors), или либеринов, а тормозящие эти процессы — ингибирующих гормонов, или статинов.
В передней части гипоталамуса находятся два ядра (паравентрикулярное и супраоптическое) с крупными нейронами, в которых синтезируются нейрогормоны окситоцин и вазопрессин. Аксоны этих нейронов формируют гипоталамо-гипофизарный тракт, по которому гормоны транспортируются из тел клеток в нейрогипофиз. Окончания аксонов образуют тесные контакты с капиллярами, в которые и выделяются гормоны. Таким образом, нейрогипофиз сохраняет и по мере необходимости выделяет в кровь гормоны, синтезированные в гипоталамусе.
Таким образом, гипоталамус на основании анализа со стояния внутренней среды организма запускает три группы реакций:
1) вегетативной нервной системы, направленные на поддержание гомеостаза;
2) поведенческие, направленные на удовлетворение потребностей организма;
3) эндокринной системы (главным образом через гипофиз).
Эпиталамус (см. рис. 32) занимает медиодорсокаудальное положение относительно других структур промежуточного мозга, т.е. находится в верхней (дорсальной) и задней (каудальной) его части посередине (медиально). Он занимает очень небольшой объем мозга и кроме различных нервных образований содержит железу внутренней секреции эпифиз (шишковидное тело).
К нервным образованиям эпиталамуса относятся поводки (или уздечки), habenula, треугольники поводков, комиссура поводков, ядра поводков.
Волокна, соединяющие эпиталамус с различными структурами переднего мозга (мозговые полоски), проходят по границе между медиальной и дорсальной сторонами таламуса. В каудальной части полоски переходят в поводок, который расширяется, образуя треугольник поводка. Медиальные части двух треугольников соединены волокнами комиссуры поводков. Под ней проходит задняя комиссура (см. 7.2.5). К комиссуре повод ков и задней комиссуре прикреплен эпифиз. В глубине треугольников поводков лежит серое вещество — ядра поводков (хабенулярные ядра), которые входят в лимбическую систему.
Функции эпиталамуса до сих пор не вполне ясны. По-видимому, в первую очередь, они связаны с деятельностью эпифиза, а нервные элементы эпиталамуса обеспечивают управление этой железой.
Эпифиз иннервируется симпатической нервной системой. Кроме того, он получает волокна от супрахиазменного ядра гипоталамуса, которое имеет прямые входы от зрительного нерва. Благодаря этому эпифиз получает информацию об уровне освещенности. Основной гормон эпифиза— мелатонин. Выяснилось, что ежедневные колебания его концентрации ритмичны и прямо связаны со световым циклом — концентрация мелатонина больше ночью. Это позволяет говорить о важной роли эпифиза в регуляции суточных ритмов. Мелатонин также влияет на половое созревание и половое поведение, тормозя активность половых желез.
Субталамус,как уже было сказано, находится на границе между средним мозгом и гипоталамусом. Структуры субталамуса можно увидеть только на разрезе мозга. Они включают несколько парных ядер серого вещества, разделенных прослойками белого вещества. Наиболее крупное ядро субталамуса — субталамическое ядро (тело Люиса).
Белое вещество включает тракты, проходящие из красного ядра среднего мозга в конечный мозг, а также собственные афференты и эфференты субталамуса. Основные афференты субталамус получает из конечного мозга — от коры больших полушарий и базальных ядер. Эфференты субталамуса идут в РФ продолговатого мозга и моста, в черную субстанцию и красное ядро (структуры среднего мозга), а также к базальным ядрам.
Из характера связей ясно, что субталамус входит в экстрапирамидную систему мозга (см. 6.4). Он принимает большое участие в организации движений, в частности локомоции — ритмических сгибаний и разгибаний конечностей и туловища, обеспечивающих перемещение тела в пространстве.
Конечный мозг
Конечный мозг, telencephalon, — наиболее массивный от дел мозга человека. Он занимает большую часть полости черепа. Конечный мозг состоит из парных больших полушарий, hemispheria cerebri, разделенных продольной щелью и прикрывающих сверху большую часть мозгового ствола и мозжечок. Выпуклая верхняя поверхность больших полушарий имеет три полюса: лобный, височный и затылочный. Нижняя поверхность больших полушарий уплощена. Длина полушария примерно 17,5 см, ширина—6,5 см. Снаружи полушария покрыты серым веществом — корой больших полушарий, ее также называют плащом или мантией. Под корой находится белое вещество, в глубине которого лежат базальные ядра (ядра конечного мозга, базальные ганглии). Полостями полушарий являются боковые желудочки.
Белое вещество полушарийсостоит из трех систем волокон:
1. Проекционные волокна представляют собой восходящие и нисходящие пути, связывающие полушария с остальными отделами ЦНС. Примером нисходящих волокон могут служить волокна кортикоспинального (пирамидного), кортикорубрального и кортикомостовых трактов, а восходящих — волокна, идущие от таламуса к коре.
2. Ассоциативные волокна связывают различные области коры одного полушария.
3. Комиссуральные волокна соединяют симметричные отделы правого и левого полушарий. Самая большая комиссура мозга — мозолистое тело, corpus callosum, представляет собой толстую горизонтальную пластинку, находящуюся в глубине продольной щели, разделяющей полушария. От этой пластинки в толще полушарий расходятся волокна, образующие лучистость мозолистого тела. В мозолистом теле выделяют (см. рис. 41) переднюю часть — колено, среднюю — тело и заднюю — валик. Колено загибается вниз и переходит в клюв мозолистого тела. Кроме мозолистого тела, в состав конечного мозга входит передняя комиссура, которая соединяет некоторые обонятельные структуры и участки височных долей, куда не распространяются каллозальные волокна (волокна мозолистого тела). Базальные ядравключают хвостатое ядро, бледный шар, скорлупу, ограду и миндалевидное тело (рис. 37, 38).
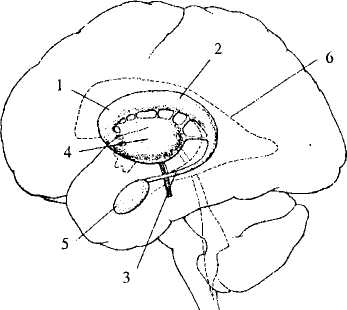
Рис. 37. Базальные ядра:
1-3 — хвостатое ядро: 1 — головка, 2 — тело, 3 — хвост;
4— скорлупа и бледный шар; 5 — миндалевидное ядро;
6 — боковой желудочек
Самым крупным из этих ядер является хвостатое ядро. Оно вытянуто в ростро-каудальном направлении (спереди назад) и имеет С-образную форму. Утолщенная передняя часть образует головку хвостатого ядра, она переходит в тело и заканчивается хвостом. На фронтальном срезе (рис. 38) видна только головка этого ядра.
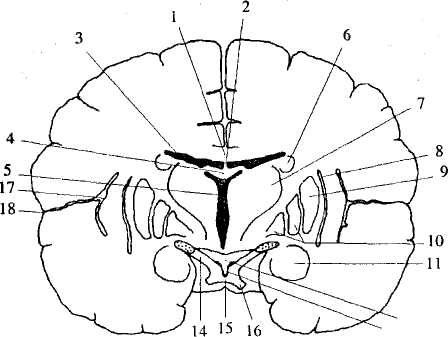
Рис. 38. Фронтальный разрез через большие полушария
на уровне серого бугра: 1 — мозолистое тело; 2 — прозрачная перегородка;
3 — центральная часть бокового желудочка; 4— свод; 5 — III желудочек;
6— хвостатое ядро; 7— таламус; 8 — ограда; 9 — скорлупа;
10— бледный шар; 11 — миндалевидное тело; 12 — серый бугор;
13— воронка; 14— зрительный тракт; 15 — зрительная хиазма;
16— зрительный нерв; 17— кора островковой доли;
18— боковая борозда
Бледный шар, скорлупа и ограда находятся латеральнее и ниже от хвостатого ядра. Они отделены от него прослойкой белого вещества (волокна корковых трактов). Самое медиальное положение занимает бледный шар, латеральнее от него лежит чашеобразная скорлупа, отделенная от бледного шара полоской белого вещества. Между скорлупой и островковой корой (см. далее) лежит полоска серого вещества — ограда.
Хвостатое ядро, бледный шар и скорлупа на разрезе выглядят как чередующиеся полоски серого и белого вещества. Из-за этого они были объединены под общим названием полосатое тело, corpus striatum. В дальнейшем при изучении клеточного состава и характера связей базальных ганглиев выясни лось, что бледный шар является филогенетически более древним образованием и значительно отличается от хвостатого ядра и скорлупы. В связи с этим бледный шар, globus pallidus, выделяют из полосатого тела как отдельную единицу — паллидум. Филогенетически более молодые хвостатое ядро и скорлупу принято называть стриатум. Вместе они образуют стриапаллидарную систему, имеющую очень обширные связи, в первую очередь, с таламусом, а также с корой больших полушарий, мозжечком, черной субстанцией, красным ядром. Очень значительные связи открыты и внутри самой системы — между отдельными ее ядрами.
Основные функции стриапаллидарной системы связаны с управлением движениями. Наряду с мозжечком, она является крупнейшим подкорковым двигательным центром. При этом если мозжечок связан с регуляцией конкретных параметров выполняемых движений (амплитудой мышечных сокращений, их согласованностью при одновременной реализации и т.п.), то стриапаллидарная система рассматривается как область, управляющая запуском движений и содержащая информацию о двигательных программах — последовательных комплексах движений. Действительно, при запуске движений активация нервных клеток наблюдается сначала в ассоциативной лобной коре, затем в стриатуме и бледном шаре, и лишь затем — в моторной коре больших полушарий и мозжечке. Как и мозжечок, структуры стриапаллидарной системы участвуют в двигательном обучении и превращении исходно произвольных (т.е. выполняемых под контролем сознания) движений в автоматизированные. При повреждении, например, стриатума наблюдается запуск патологических движений — высокоамплитудных подергиваний рук (хорея), скручиваний туловища (атетоз). Проявления паркинсонизма (тремор и т.п.) также связаны в основном с нарушением влияния черной субстанции на хвостатое ядро.
Миндалевидное тело, corpus amygdaloideum, — сферическое образование, располагающееся под скорлупой около внутренней части переднего отдела височной коры. Амигдала (миндалина) соприкасается с хвостом хвостатого ядра, который, закручиваясь, заходит в височные доли. Она имеет многочисленные связи с корой больших полушарий, гипоталамусом, обонятельными мозговыми структурами. Амигдала входит в лимбическую систему мозга и играет большую роль в организации эмоций. Повреждение миндалины часто ведет к глубоким изменениям психики, депрессивным и маниакальным состояниям.
Кора больших полушарий— высший отдел ЦНС, она отвечает за восприятие всей поступающей в мозг информации, за управление сложными движениями, мыслительную и речевую деятельность. Филогенетически это самое молодое образование нервной системы. Впервые в эволюции она появляется у пресмыкающихся, но в полном объеме развивается только у млекопитающих.
Кора больших полушарий человека и ряда других млеко питающих имеет складчатый вид. На ее поверхности выделяют многочисленные извилины, разделенные бороздами, что очень увеличивает ее площадь. Поверхность коры обоих полушарий взрослого человека колеблется от 1470 до 1670 см2. Крупные борозды разделяют каждое полушарие на пять долей — лобную, теменную, затылочную, височную и островок. Островок, insula, — доля, не выходящая на поверхность полушария; инсулярная кора расположена в глубине латеральной борозды, представляет собой расширение ее дна и при крыта височной долей (см. рис. 38). Кроме этого, в коре можно выделить лимбическую долю, расположенную на медиальной (срединной) поверхности и представляющую собой группу извилин, окружающих ствол мозга и мозолистое тело (см. рис. 41). Для человека характерно преобладание лобной и височной долей, поверхность которых в сумме составляет 47% от всей поверхности полушарий.
Основные борозды и извилины коры больших полушарий представлены на рис 39, 40, 41.
Расположение борозд и извилин на латеральной (боковой) поверхности (рис. 39) не представляет сложностей для изучения. Отметим лишь, что здесь находятся две самые глубокие борозды — центральная (роландова), отделяющая лобную долю от теменной, и латеральная (боковая или сильвиева), отделяющая височную долю от лобной и теменной. Перед центральной бороздой лежит передняя центральная (прецентральная) извилина, а за ней — задняя центральная (постцентральная) извилина. Теменная доля отделяется от затылочной теменно-затылочной бороздой, отчетливо видной только на медиальной поверхности мозга (см. рис. 41)
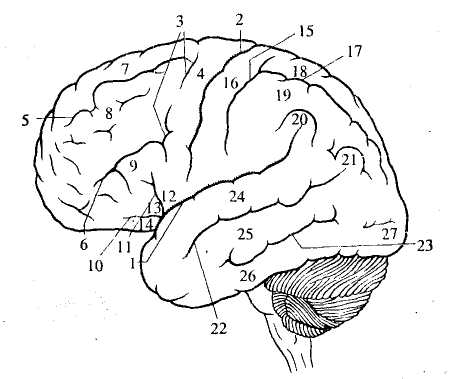
Рис. 39. Латеральная поверхность полушарий:
1 — боковая (сильвиева) борозда; 2 — центральная (роландова)
борозда; 3 - 14— лобная доля: 3 — прецентральная борозда,
4— прецентральная извилина, 5 — верхняя лобная борозда,
6 — нижняя лобная борозда, 7 — верхняя лобная извилина,
8 — средняя лобная извилина, 9 — нижняя лобная извилина,
10— передняя ветвь, 11 — восходящая ветвь, 12 — покрышечная
часть, 13 — треугольная часть, 14 — глазничная (орбитальная)
часть; 15 -21 — теменная доля: 15 — постцентральная борозда,
16— постцентральная извилина, 17— внутритеменная борозда,
18— верхняя теменная долька, 19 — нижняя теменная долька,
20 — надкраевая извилина, 21 — угловая извилина; 22-26 — височная
доля: 22 — верхняя височная борозда, 23 — нижняя височная борозда,
24— верхняя височная извилина, 25 — средняя височная извилина,
26— нижняя височная извилина; 27 — затылочный полюс
Рассматривая базальную (нижнюю) поверхность полушарий (рис. 40), нужно иметь в виду, что ствол мозга на этом рисунке удален (сравните с рис. 21 — нижняя поверхность головного мозга). Нижняя поверхность лобной доли носит название «орбитальная кора».
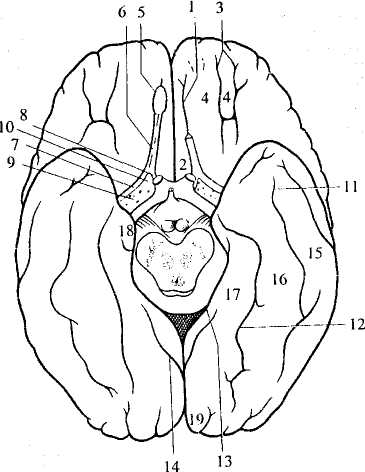
Рис. 40. Нижняя поверхность полушарий:
1 – 10 — лобная доля: 1 — обонятельная борозда, 2 — прямая извилина,
3— глазничные борозды, 4— глазничные извилины, 5 — обонятельная
луковица, 6— обонятельный тракт, 7—латеральная обонятельная
полоска, 8 — медиальная обонятельная полоска, 9 — передняя
продырявленная субстанция, 10 — обонятельный треугольник;
11 – 19 — височная и затылочная доли: 11 — затылочно-височная
борозда, 12 — окольная борозда, 13 — гиппокампальная борозда,
14— шпорная борозда, 15— латеральная затылочно-височная
извилина, 16 — медиальная затылочно-височная извилина,
17— парагиппокампальная извилина, 18 — крючок,
19— язычная извилина
Справа обонятельная луковица и часть обонятельного тракта удалены.
В медиальной части лобной доли проходит обонятельная борозда, в которой лежит обонятельная луковица и идущий от нее обонятельный тракт (на рисунке в левом полушарии они удалены). К нижней поверхности обонятельной луковицы подходят волокна обонятельного нерва. Обонятельный тракт в своем основании разветвляется на латеральную, среднюю и медиальную обонятельные полоски. Между латеральной и медиальной полосками лежит обонятельный треугольник. В его глубине находится переднее обонятельное ядро, а в его основании — переднее продырявленное вещество. Через него, как и через заднее продырявленное вещество (см. 7.2.5), в мозг входит много сосудов.
Большинство борозд и извилин затылочной и височной долей видны как на нижней, так и на медиальной поверхности полушарий (см. рис. 40, 41). Это затылочно-теменная, окольная, гиппокампальная и шпорная борозды; медиальная и латеральная затылочно-теменные извилины, язычная и парагиппокампальная извилины, а также извилина, называемая крючок.
Наибольшее число образований переднего мозга видно на медиальной {срединной) поверхности (рис. 41). Для того что бы лучше представить себе взаимное пространственное рас положение структур, полезно сравнить этот рисунок с рис. 38 (горизонтальный разрез через базальные ганглии) и рис. 20 (медиальный разрез через головной мозг).
В центре медиальной поверхности находится мозолистое тело. В основании его клюва видна передняя комиссура. Под мозолистым телом проходят волокна свода, идущие от гиппокампа к мамиллярным телам. Между сводом и коленом мозолистого тела натянута тонкая пластинка из глиальных клеток — прозрачная перегородка. Между перегородками правого и левого полушарий есть небольшая полость, которую рассматривают иногда как V мозговой желудочек. Рядом с перегородкой (под ее нижней частью) находятся скопления нейронов — ядра прозрачной перегородки, входящие в лимбическую систему.
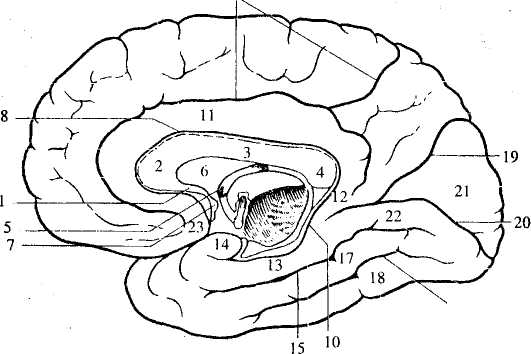
Рис. 41. Медиальная поверхность полушарий:
1—4 — мозолистое тело: 1 — клюв, 2 — колено, 3 — тело, 4 — валик',
5— свод; 6— прозрачная перегородка; 7— передняя комиссура;
8— борозда мозолистого тела; 9 — поясная борозда; 10 — гиппокампальная борозда; 11 — поясная извилина; 12 — перешеек;
13 — парагиппокампальная извилина; 14 — крючок; 15 — окольная
борозда; 16— затылочно-височная борозда; 17 — медиальная
затылочно-височная извилина; 18 — латеральная затылочно-височная
извилина; 19— теменно-затылочная борозда; 20— шпорная борозда;
21 — клин; 22— язычная извилина; 23 — подмозолистая извилина
Между мозолистым телом и корой проходит борозда мозолистого тела. Над ней расположена поясная борозда. Она начинается под клювом мозолистого тела, а ее задний конец загибается вверх. Между этими бороздами лежит поясная извилина. Сзади она продолжается в перешеек, который снизу переходит в парагиппокампальную извилину, заканчивающуюся крючком. Парагиппокампальная извилина ограничена гиппокампальной и окольной бороздами. Вместе поясная извилина, перешеек, парагиппокампальная извилина и крючок обозначаются как сводчатая извилина. Она описывает почти полный круг и рассматривается как лимбическая доля полушария. В глубине гиппокампальной борозды, практически не видная на поверхности мозга, лежит зубчатая извилина.
В лобной доле отметим подмозолистую извилину, расположенную перед клювом мозолистого тела.
В задней части полушария проходит теменно-затылочная борозда, отделяющая теменную долю от затылочной. Участок между ней и шпорной бороздой называется клином.
По своему филогенезу кора больших полушарий разделяется на древнюю, paleocortex, старую, archicortex, и новую, neocortex. Большую часть коры (у человека 96%) занимает новая кора. Древняя и старая кора (палеокортекс и архикортекс) занимает лишь небольшие участки на базальной и медиальной поверхностях лобной и височной долей полушарий. Эти участки более примитивны по своей структуре. Новая кора (неокортекс) имеет шестислойное строение, которое не является характерным для древней и старой коры. Большинство структур палео- и архикортекса входят в лимбическую систему мозга.
Палеокортекс включает структуры, связанные главным образом с анализом обонятельных раздражителей: обонятельные луковицы, обонятельный тракт, обонятельные треугольники, передние обонятельные ядра, подмозолистая извилина, септальная область (подмозолистая извилина и ядра перегородки — скопления серого вещества под клювом мозолистого тела), передние участки медиальной поверхности височных долей и др.
Волокна обонятельного нерва начинаются от рецепторов, находящихся в стенке носовой полости. Заканчиваются они в обонятельной луковице, находящейся на нижней поверхности лобной доли больших полушарий. Аксоны нейронов обонятельной луковицы образуют обонятельный тракт, волокна которого идут в перечисленные выше структуры, а также в ряд других образований, например в гипоталамус.
Большинство структур палеокортекса входит в лимбическую систему, в связи с чем можно говорить о его участии в организации эмоционального поведения.
Архикортекс.Это морской конек, илигиппокамп, hippocampus, находящийся на внутренней поверхности височной доли, и зубчатая извилина. Гиппокамп тянется вдоль всей медиальной стенки нижнего рога бокового желудочка. Он образуется в результате того, что в процессе онтогенеза гиппокампальная борозда впячивается в стенку нижнего рога, вдавливая туда мозговое вещество. Таким образом, гиппокамп расположен на дне гиппокампальной борозды за парагиппокампальной извилиной. Зубчатая извилина, которая уже упоминалась при описании медиальной поверхности полушарий, находится несколько медиальнее гиппокампа. Через зубчатую извилину проводится информация от различных корковых зон к гиппокампу.
В переводе с латинского, hippocampus — морской конек. Он назван так за характерную форму своего поперечного разреза.
Как уже упоминалось, от гиппокампа к мамиллярным те лам гипоталамуса идет толстый пучок волокон — свод. Свод включает аксоны, идущие от каждого из гиппокампов — ножки свода (между ними проходит комиссура). Далее, огибая таламус, ножки объединяются в тело свода. При подходе к мамиллярным телам они снова расходятся, образуя столбы свода (рис. 42). Небольшая часть волокон свода идет в другие образования (таламус, миндалину, структуры палеокортекса).
Гиппокамп тесно связан с процессами научения и памяти. При различных повреждениях гиппокампа нарушаются процессы запоминания.
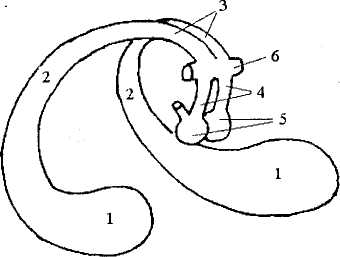
Рис. 42. Гиппокамп и свод:
1 — гиппокамп; 2 — ножки свода; 3 — тело свода;
4 — столбы свода; 5— мамиллярные тела; 6— передняя комиссура
Неокортекс.Новая кора имеет шесть слоев (рис. 43), ее толщина у человека примерно 3 мм.
В каждом корковом слое преобладают нейроны определенного строения. Различаются и функции этих нейронов. Аксоны нервных клеток коры и подходящие к ним волокна из других структур образуют белое вещество полушарий.
С системой нисходящих проекционных волокон главным образом связан 5-й слой (если считать от поверхности)— внутренний пирамидный слой. В нем находятся крупные клетки, тела которых имеют форму пирамид (см. рис. 9, Г). Аксоны этих клеток образуют главный эфферентный путь коры больших полушарий — кортикоспинальный (пирамидный). Именно в пирамидных нейронах генерируются им пульсы, управляющие в конечном итоге работой мышц при произвольных движениях. Основные афферентные волокна, поступающие в кору из таламуса, оканчиваются на нейронах 4-го (внутреннего зернистого) слоя, в который входят мелкие зернистые и звездчатые нейроны.
Каллозальные и ассоциативные волокна главным образом идут от нейронов 3-го слоя (наружного пирамидного), а приходят во 2-й слой (наружный зернистый).
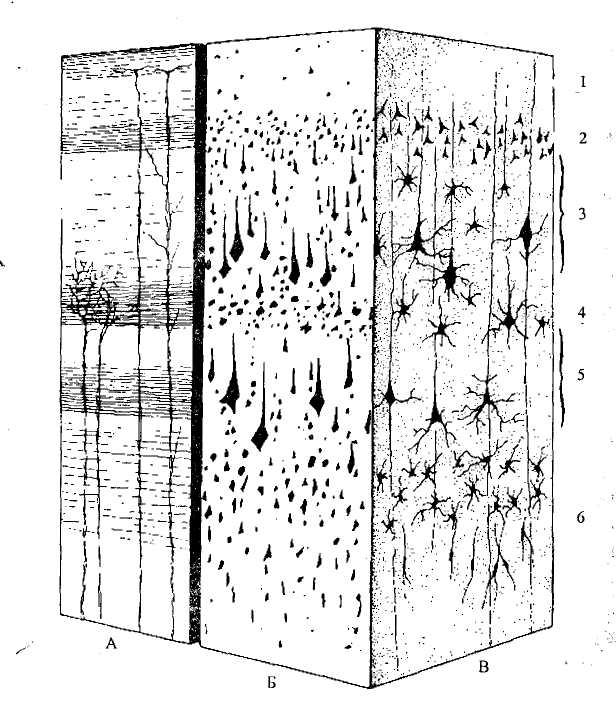
Рис. 43. Слои неокортекса:
А, Б, В — изображение коры при различных видах окраски (А — окрашены только отростки нейронов, Б — окрашены только тела нейронов, В — нейроны окрашены целиком).
1 - молекулярный слой; 2 — наружный зернистый слой; 3 — наружный пирамидный слой; 4 — внутренний зернистый слой; 5 — внутренний пирамидный слой; 6 — полиморфный слой
Кора больших полушарий дополнительно подразделяется на 52 поля, отличающиеся по своему клеточному строению и функциям (рис. 44).
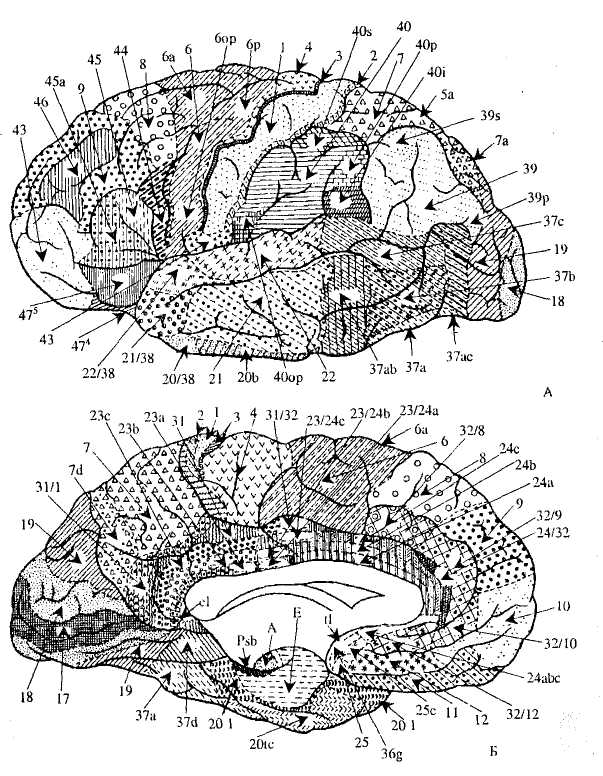
Рис. 44. Поля коры больших полушарий мозга человека: А — латеральная поверхность, Б — медиальная поверхность
В настоящее время кору больших полушарий принято разделять на сенсорные (первичные проекционные), двигательные и ассоциативные зоны (рис. 45).
К сенсорным зонам коры относят поля, в которые приходят волокна от проекционных ядер таламуса. Это зоны коркового представительства сенсорных систем. Здесь заканчиваются самые короткие пути от рецепторов. Для этих зон характерно очень сильное развитие 4-го слоя коры и в то же время плохо выраженный 5-й слой.
Для каждой сенсорной системы существуют свои проекционные зоны. Зрительная зона находится в затылочной доле коры больших полушарий. Она расположена на участке под названием «клин» на медиальной поверхности. Слуховая зона расположена в верхней височной извилине. Вкусовые зоны найдены в нижней части постцентральной извилины и в инсулярной (островковой) коре. Корковые обонятельные зоны уже были описаны (см. палеокортекс).
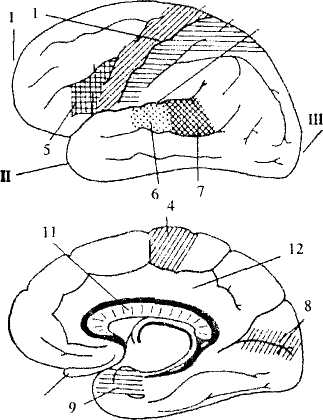
Рис. 45. Основные зоны коры больших полушарий
Вверху — боковая поверхность левого полушария.
Внизу — срединная поверхность правого полушария.
I — лобный полюс, II — височный полюс, III — затылочный полюс,
1- центральная борозда; 2 - боковая борозда; 3 - зона кожной
чувствительности; 4 — двигательная зона; 5 — центр Брока; 6 — зона
слуховой чувствительности; 7 — центр Вернике; 8 - зона зрительной
чувствительности; 9— зона обонятельной чувствительности;
10 - обонятельная луковица; 11 -мозолистое тело;
12-лимбическая доля
Большой участок занимает зона кожно-мышечной чувствительности. Она расположена позади центральной борозды, в постцентральной извилине теменной доли коры больших полушарий. Как уже отмечалось (см. 6.4), кожно-мышечные проекции организованы по соматотопическому принципу. Но «карта тела» в коре имеет несколько смещенные пропорции. Дело в том, что количество нейронов, получающих информацию от определенного участка тела, прямо пропорционально плотности рецепторов на этом участке. Плотность же рецепторов зависит от значимости информации, получаемой от того или иного участка кожной поверхности. Поэтому на «карте тела» в коре отмечаются непропорционально большие зоны кисти рук и губ, но очень маленькие зоны спины, живота и т.д. (рис. 46).
Двигательная (моторная) зона расположена в прецентральной извилине лобной доли коры больших полушарий впереди центральной борозды. Именно отсюда отходит большинство волокон кортикоспинального тракта. Как уже говорилось, этот путь начинается в 5-м слое коры, который выражен здесь значительно сильнее, чем в остальных зонах. В то же время 4-й слой в моторной коре практически отсутствует, за что кора в этой области получила название «агранулярная» (гранулярный — зернистый). Так же, как и в зоне кожно-мышечной чувствительности, в прецентральной извилине можно нарисовать «карту тела», причем она также будет иметь определенные искажения пропорций человеческого тела (рис. 46). Связано это с тем, что некоторые мышцы (пальцев, мимические) должны выполнять гораздо более тонкие движения, поэтому для управления ими необходимо большее количество нейронов.
К неспецифичной или ассоциативной коре отнесены области, которым нельзя приписать каких-либо преимущественно сенсорных или двигательных функций. У человека ассоциативные зоны занимают больше половины всей коры. Эти зоны связывают (ассоциируют) друг с другом сенсорные и двигательные зоны и одновременно служат субстратам высших психических функций.
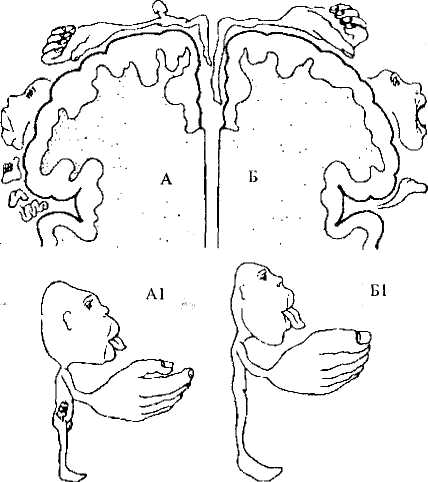
Рис. 46. Представительство зон кожной чувствительности
и двигательных зон в коре больших полушарий. Проекционная зона
кожной чувствительности (А). Моторные зоны (Б).
Несоразмерность пропорций проиллюстрирована в виде сенсорного (А1) и моторного (Б1) гомункулюса (человечка)
Основные неспецифические области коры больших полушарий — это теменно-височно-затылочная (находится на границе этих долей), префронтальная (лобная доля коры за исключением прецентральной извилины) и лимбическая (сводчатая извилина) ассоциативные зоны. Если упрощенно представить их функции, каждая из этих областей особенно важна соответственно для следующих интегративных процессов: высших сенсорных функций и речи; высших двигательных функций и запуска поведенческих актов; памяти и эмоционального поведения.
Хотя по своему строению правое и левое полушария чело века практически не отличаются, для них характерна функциональная асимметрия, т.е. они выполняют разные функции. В первую очередь это относится к ассоциативным зонам коры. В повседневной жизни эти различия не заметны, так как ин формация легко переходит из полушария в полушарие через комиссуры мозга (в первую очередь через мозолистое тело). Представления о различиях в функциях полушарий сложились при наблюдениях за больными с односторонними поражения ми мозга и в специальных экспериментах, где информация поступала только в одно из полушарий.
Оказалось, что левое полушарие (по крайней мере у правшей) в большей степени связано с речью, абстрактно-понятийным мышлением, математическими способностями. Правое полушарие преимущественно управляет образным мышлением и в значительной мере определяет такие свойства, как музыкальность, распознавание сложных зрительных образов, выражение и восприятие эмоций.
В лобной и височной областях левого полушария находятся центры речи (см. рис. 45). При поражении центра речи в височной коре (центр Вёрнике), находящегося на границе со слуховой корой, нарушается понимание слышимой речи. При поражении же центра речи, лежащего на границе сдвигательной областью в лобной коре (центр Брока), больной слышит и понимает речь, но сам говорить не может. При поражении не которых областей правого полушария отмечаются глубокие нарушения ориентации в пространстве, например, больные не могут найти дорогу к дому, в котором прожили много лет. Не которые пациенты с повреждениями правого полушария не могут узнавать знакомые лица, а иногда вообще не могут узнавать людей.
В заключение необходимо подчеркнуть, что мозг обладает чрезвычайно большими компенсаторными возможностями. Конечно, многим его зонам (ядрам) присущиврожденно обусловленные функции (особенно это характерно для первичных сенсорных зон, центров продолговатого мозга и т.п.). Однако очень многие области, в первую очередь, корковые образования, приобретают конкретные «обязанности» по мере созревания и обучения ЦНС. Это свойство мозга предопределяет возможность того, что при поражении различных участков соответствующие функции принимают на себя другие отделы ЦНС.
8. Вегетативная (автономная)
нервная система
Все функции организма можно разделить на соматические, «животные», и вегетативные, «растительные». Соматические функции связаны с восприятием внешних раздражений и двигательными реакциями, осуществляемыми скелетной мускулатурой. От вегетативных функций зависит осуществление обмена веществ в организме (пищеварение, кровообращение, дыхание, выделение и т.д.), а также рост и размножение.
Как известно (см. 1.2), помимо морфологических различий между гладкой и скелетной мускулатурой существуют и функциональные отличия между ними. Скелетная мускула тура отвечает на воздействие среды быстрыми целесообразными движениями, которые можно регулировать произвольно. Гладкая мускулатура, заложенная во внутренних органах и сосудах, работает медленно, но ритмично; ее деятельность обычно не поддается произвольной регуляции. Эти функциональные различия связаны с разницей в иннервации: скелетные мышцы получают импульсы от соматической части НС, гладкие — от вегетативной. Вегетативная нервная система (ВНС) иннервирует не только гладкую мускулатуру, но и другие неподдающиеся произвольной регуляции исполнительные органы — сердечную мышцу и железы[2].
В целом ВНС выполняет адаптационно-трофическую функцию, т.е. приспосабливает уровень активности тканей и органов к выполняемым ими в текущий момент времени задачам. Эти задачи в свою очередь обычно связаны с той или иной деятельностью организма в изменяющихся условиях внешней среды.
Дуги вегетативных рефлексов и их отличия от дуг соматических рефлексов были рассмотрены ранее (см. 6.2, рис. 19, Б).
Напомним, что в автономной нервной системе эфферентная часть дуги состоит из двух нейронов: преганглионарного (последнего или единственного центрального нейрона) и ганглионарного (расположенного в вегетативном ганглии). Из такого расположения нейронов следует главный признак ВНС — двухнейронность эфферентного пути.
Аксоны центральных нейронов ВНС, которые заканчиваются на клетках вегетативных ганглиев, называют преганглионарными волокнами, а аксоны исполнительных нейронов (которые расположены в ганглиях) — постганглионарными. Преганглионарные волокна покрыты миелиновой оболочкой, постганглионарные отличаются ее отсутствием (так называемые серые волокна).
Из правила двухнейронности эффекторного пути имеются некоторые исключения:
1. Постганглионарные симпатические волокна, идущие к гладким мышцам желудочно-кишечного тракта, преимущественно оканчиваются не на мышечных волокнах, а на парасимпатических ганглионарных клетках, находящихся в стенках желудка и кишок. По-видимому, они снижают активность этих клеток и таким образом оказывают тормозящее влияние на гладкую мускулатуру (3-нейронная структура эфферентного пути).
2. Хромаффинные клетки мозгового слоя надпочечников иннервированы не пост-, а преганглионарными симпатическими волокнами. Хромаффинные клетки образуют под влиянием импульсов, поступающих к ним по симпатическим волокнам, адреналин. Эти клетки по своей сути соответствуют постганглионарным нейронам, с которыми имеют общее происхождение из ганглиозной пластинки (1-нейронная структура эфферентного пути).
ВНС делится на два отдела — симпатический и парасимпатический, которые принято называть системами. Большинство органов иннервируется как симпатическими, так и парасимпатическими волокнами. Однако в некоторых случаях наблюдается преобладающее значение какого-либо отдела. Слезные железы и железы носоглотки иннервируются только парасимпатической НС. В основном парасимпатическую ин нервацию имеет мочевой пузырь. С другой стороны, только симпатические волокна подходят к гладким мышцам кровеносных сосудов (за исключением сосудов мозга и артерий половых органов), потовых желез, селезенки, секреторным клеткам надпочечников.
Симпатическая и парасимпатическая системы отличаются друг от друга функционально (по выполняемой деятельности), морфологически (по строению), а также медиаторами, используемыми при передаче нервного импульса.
Функциональные отличиясвязаны с тем, что симпатическая и парасимпатическая системы, как правило, противоположным образом влияют на различные органы и ткани. Если симпатический отдел возбуждает какую-либо часть организма, то парасимпатический тормозит ее и наоборот. Так, раздражение симпатического нерва, иннервирующего сердце, усиливает его работу, а раздражение парасимпатического блуждающего нерва тормозит сердечные сокращения. Одна ко не следует думать, что между симпатической и парасимпатической частями ВНС существует жесткий антагонизм, а их функции полностью противопоставлены. Это части взаимодействующие, соотношение между ними динамически меняется на разных фазах деятельности того или иного органа, т.е. они функционируют согласованно. Например, и симпатическая, и парасимпатическая стимуляция вызывает выделение слюны. Но в первом случае слюна будет густой, насыщенной органическими веществами, а во втором — жидкой, водянистой.
В регуляции активности всей ВНС участвуют гипоталамус (высший вегетативный центр), ретикулярная формация, ряд других вегетативных центров.
Симпатическая нервная системаподготавливает организм к активным действиям. Она увеличивает обмен веществ, усиливает дыхание и работу сердца, увеличивает поступление кислорода к мышцам, расширяет зрачок, тормозит работу пищеварительной системы, сокращает сфинктеры (круговые запирательные мышцы) некоторых полых органов (мочевого пузыря, желудочно-кишечного тракта), расширяет бронхи. Работа симпатической нервной системы усиливается при стрессогенных раздражителях.
Парасимпатическая нервная системавыполняет охранительную функцию, она способствует расслаблению организма и восстановлению его энергетических запасов. Раздражение парасимпатических волокон приводит к ослаблению работы сердца, сокращению зрачка, усилению моторной и секреторной деятельности желудочного - кишечного тракта, опорожнению полых органов, сужению бронхов.
Таким образом, симпатический отдел нервной системы приспосабливает организм к интенсивной деятельности. Парасимпатический отдел нервной системы способствует восстановлению истраченных ресурсов организма. Каждый из них имеет центральную и периферическую части.
Морфологические отличиямежду симпатической и парасимпатической системами связаны с местонахождением последних двух нейронов (центрального и периферического) дуги вегетативного рефлекса (рис. 47). Такие нейроны сгруппированы в вегетативные ядра внутри ЦНС и в вегетативные ганглии в периферической НС. Симпатические ядра расположены в грудном и верхнем поясничном отделах спинного мозга (в боковых его рогах), а парасимпатические ядра — в стволе головного мозга и крестцовом отделе спинного мозга (в промежуточном веществе). В связи с положением центральных нейронов симпатическую систему принято называть грудинно-поясничной, или торако-люмбальной (thoracale — грудной; lumbale — поясничный), а парасимпатическую — черепно-крестцовой, или кранио-сакральной (kranion — череп; sacrale — крестцовый).
А Б
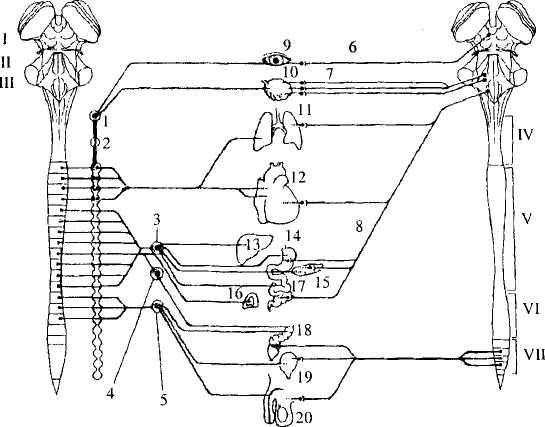
Рис. 47. Разделение вегетативной нервной системы на симпатическую (А) и парасимпатическую (Б) части: I – VII — структуры ЦНС: I — средний мозг, II — мост, III — продолговатый мозг, IV — шейный, V — грудной, VI — поясничный, VII — крестцовый отделы спинного мозга; 1-8— структуры периферической нервной системы: 1 — верхний шейный ганглий, 2— звездчатый ганглий, 3 — солнечное сплетение, 4 — верхний брыжеечный ганглий, 5 — нижний брыжеечный ганглий, 6 — волокна глазодвигательного нерва, 7 — волокна лицевого и языкоглоточного нерва, 8 — волокна блуждающего нерва; 9 — 20 — внутренние органы: 9— глаз, 10— слезные и слюнные железы, 11 — легкие,
12— сердце, 13— печень, 14— желудок, 15 — поджелудочная
железа, 16— надпочечники, 17 — кишечник, 18 — прямая кишка,
19— мочевой пузырь, 20— половые органы
Симпатические ганглии идут вдоль позвоночника, образуя две (правую и левую) симпатические цепочки. В цепочках выделяют шейный, грудной, поясничный и крестцовый отделы, в каждом из которых имеется несколько пар ганглиев. Надо отметить, что в рефлекторной дуге симпатической НС последний нейрон может находиться не только в узлах симпатического ствола, но и в нервных сплетениях (ganglia celiaca— чревный узел, g.mesenterica — брыжеечный узел и др.). Парасимпатические ганглии находятся или рядом с иннервируемым органом (экстрамуральные ганглии), или в его стенках (интрамуральные ганглии). Таким образом, получается, что преганглионарные волокна симпатической нервной системы короткие, а постганглионарные — длинные. Для парасимпатической системы характерна обратная закономерность.
Необходимо отметить, что число нервных клеток в ганглиях в несколько раз больше числа преганглионарных волокон (в шейном симпатическом узле — в 32 раза, в ресничном узле — в 2 раза). Соответственно, каждое из преганглионарных волокон ветвится и образует синапсы на нескольких клетках ганглия. Таким образом, достигается расширение зоны влияния преганглионарных волокон.
Из изложенного ясно, что в головном мозгу нет симпатических центров. Тем не менее, гладкие мышцы и железы головы имеют симпатическую иннервацию. К этим органам подходят волокна, идущие от верхних шейных ганглиев. Они проникают в полость черепа, образуя сплетения вокруг внутренней сонной и позвоночной артерий. Кроме головы, шейные ганглии вместе с грудными иннервируют органы шеи и грудной полости. Поясничные ганглии посылают волокна к органам брюшной полости, а крестцовые — к прямой кишке и половым органам.
Парасимпатические волокна краниального отдела проходят в составе глазодвигательного, лицевого, языкоглоточного и блуждающего нервов (см. 7.2.1). Напомним, что парасимпатические волокна блуждающего нерва, выходя из полости черепа, образуют синапсы на парасимпатических ганглиях, иннервирующих большинство внутренних органов тела. Волокна, отходящие от сакрального отдела, связаны с парасимпатическими влияниями на прямую кишку, мочевой пузырь, половые органы.
Еще одно отличие между симпатической и парасимпатической системами можно назвать нейрохимическим, в связи с разными медиаторами, принимающими участие в передаче нервного импульса в ВНС. Все нейроны вегетативных ядер (т.е. центральные нейроны) — ацетилхолинергические. Таким образом, медиатор, передающий нервный импульс в вегетативных ганглиях (как симпатических, так и парасимпатических), — ацетилхолин. В то же время нейроны вегетативных ганглиев отличаются по вырабатываемому ими медиатору. В симпатической нервной системе это обычно норадреналин, а в парасимпатической — ацетилхолин. Таким образом, в симпатической нервной системе на исполнительный орган сигнал передается с помощью норадреналина, а в парасимпатической — ацетилхолина.
В последнее время в вегетативной нервной системе выделяют еще один отдел — метасимпатическую (энтеральную) нервную систему. Отличительной ее особенностью являются рефлекторные дуги, не проходящие через ЦНС. То есть и чувствительный, и вставочный, и исполнительный нейроны находятся за пределами ЦНС, непосредственно в стенках иннервируемого органа. Благодаря этому многие внутренние органы после перерезки симпатических и парасимпатических путей или даже после извлечения из организма (при создании соответствующих условий) продолжают осуществлять присущие им функции без особых видимых изменений. Например, сохраняется перистальтическая функция кишечника, сокращается промываемое физиологическим раствором сердце, сжимаются и разжимаются лимфатические сосуды и т.п.
Вместе с тем, обладая достаточно большой самостоятельностью, метасимпатическая нервная система сохраняет связь с остальной нервной системой, так как на ее нервных клетках образуют синапсы как симпатические, так и парасимпатические нейроны.
9. Лимбическая система
Лимбическая система (ЛС) — комплекс анатомически и функционально связанных между собой структур, участвующих в регуляции поведенческих реакций, в первую очередь эмоций, а также играющих значительную роль в регуляции вегетативных реакций.
Основой ЛС является так называемый круг Пейпеца, описанный еще в 1937 г. Он включает гиппокамп, идущий от него свод, затем мамиллярные тела гипоталамуса, лимбические (передние) ядра таламуса и поясную извилину (лимбическая кора). Все структуры круга Пейпеца замкнуты в кольцевую систему (рис. 48). В дальнейшем этот круг был дополнен рядом других структур. В настоящее время в ЛС включают всю лимбическую долю коры больших полушарий, архикортекс, ряд структур палеокортекса, а также некоторые подкорковые образования — миндалевидный комплекс, септальные ядра (ядра прозрачной перегородки), эпиталамус, некоторые образования среднего мозга (ретикулярная формация). Части ЛС объединены многочисленными двусторонними связями. Выявлены и связи ЛС с другими мозговыми образованиями (с базальными ганглиями, неокортексом и др.).
Необходимым условием для включения какой-либо структуры в ЛС является участие в организации мотивационно-эмоционалыюго поведения. При поражении или стимуляции различных участков ЛС наблюдаются разнообразные поведенческие, в том числе эмоциональные реакции. Так, уже упоминавшиеся (см. 7.4.1) центры положительного и отрицательного подкрепления найдены не только в гипоталамусе, но и в других структурах ЛС (поясная извилина, септальная область, амигдала).
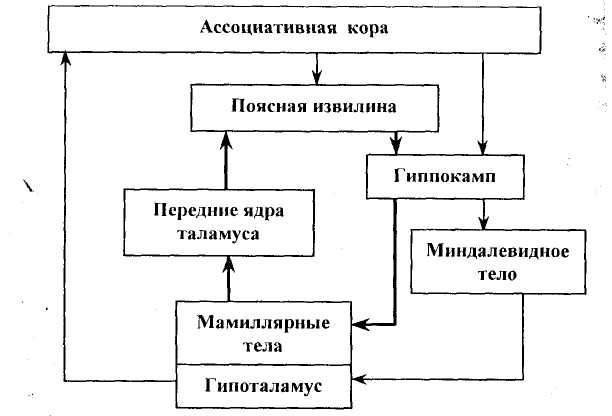
Рис. 48. Основные структуры
лимбической системы и связи между ними
(полужирными стрелками выделен круг Пейпеца)
Кроме того, все лимбические образования прямо или опосредовано связаны с гипоталамусом, высшим вегетативным центром (см. 7.4.1). В связи с последним фактом говорят об участии ЛС в регуляции вегетативных функций.
ЛС участвует в возникновении некоторых заболеваний. Это, например, отчетливо проявляется при эпилепсии. Так, локализация эпилептического очага вблизи миндалевидного тела характеризуется наличием эмоционально-мотивационных расстройств, среди которых наиболее частыми являются вспышки немотивированной агрессии, а также эмоциональные дисфории — резкие беспричинные перепады настроения. Нередко такие эмоциональные нарушения сопровождаются слуховыми, зрительными и кинестетическими галлюцинациями, отражая сложную картину поздней стадии эпилепсии, когда эпилептогенный процесс может з
Дата добавления: 2016-01-03; просмотров: 2016;