Некоторые белки человека и животных и их функции
| Тип белка | Функция |
| Структурные белки: | |
| Гликопротеиды | Образуют клеточные стенки |
| Белки мембран | Образуют мембраны |
| Белки вирусного капсида | Образуют вирусный капсид |
| Коллаген | Образуют фиброзную соединительную ткань |
| Эластин | Связки |
| Кератин | Кожа, перья, ногти, копыта |
| Ферменты | Катализируют реакции синтеза и распада |
| Гормоны: | |
| Инсулин | Регулирует обмен глюкозы |
| Аденокортикотропный | Регулирует синтез кортикостероидов |
| Гормон роста | Стимулирует рост костей |
| Двигательные белки: | |
| Миозин | Нити в миофибриллах |
| Актин | Движущиеся нити в миофибриллах |
| Дипеин | Движение жгутиков и ресничек простейших |
| Транспортные белки: | |
| Гемоглобин | Переносит кислород (позвоночные) |
| Гемоцианин | Переносит кислород (беспозвоночные) |
| Миоглобин | Переносит кислород в мышцах |
| Защитные белки: | |
| Антитела | Контролируют чужеродные белки |
| Комплемент | Комплексируются с антителом + антигеном |
| Фибриноген | Предшественник фибрина |
| Тромбин | Фактор свертываемости крови |
| Токсины: | |
| Ботулистический токсин | Пищевые отравления |
| Змеиный яд | Гидролиз фосфоглицеридов |
| Рицин | Токсический белок клещевины |
Многие белки являются ферментами (энзимами). Ферменты локализуются в митохондриях, цитоплазме, лизосомах, пероксисо-мах, на мембранах клеток и органелл. Они катализируют все протекающие в клетках реакции. Считают, что ферменты повышают скорость реакции минимум в 1 млн раз. Каждая реакция обеспечивается собственным ферментом. Например, липаза расщепляет жиры, амилаза расщепляет крахмал. В настоящее время известно более 2000 разных ферментов. В зависимости от катализируемых реакций их классифицируют на гидролазы (реакции гидролиза), нуклеазы (расщепление нуклеиновых кислот), трансферазы (перенос функциональных групп), оксидоредуктазы (окислительно-восстановительные реакции), липазы (образование связей за счет АТФ) и др.
Важнейшей особенностью белков является то, что в клетках многоклеточных организмов тысячи белков функционально связаны между собой и переносят информацию от плазматической мембраны к геному. Например; фермент в метаболическом пути «читает» концентрацию субстрата и продуцирует соответствующий уровень продукта, а рецептор на клеточной поверхности «читает» концентрацию его лиганда и продуцирует определенный уровень комплекса рецептор — лиганд.
Белки обладают регуляторной способностью. Те животные белки, которые являются гормонами, обладают способностью регулировать физиологические процессы, протекающие в клетках. Например, инсулин, являющийся белковым гормоном, продуцируемым клетками поджелудочной железы, регулирует в организме метаболизм глюкозы. Белковыми являются также гормоны, продуцируемые клетками гипоталамической части мозга и гипофизом и имеющие важное значение в росте и развитии организмов. Пара-тиреоидный гормон регулирует транспорт ионов Са^ и фосфатов. Однако заметим, что не все гормоны имеют белковую природу. У растений также известны отдельные белки, обладающие гормональной активностью. Например, такой активностью обладает индоли-луксусная кислота, которая стимулирует рост растений. Репрессорные белки участвуют в регуляции экспрессии генов.
Белки обладают двигательной и сократительной функциями, обеспечивая на молекулярном, уровне движение хромосом и сперматозоидов, на других уровнях — движение простейших, двигательные реакции у растений; сокращение скелетных мышц у многоклеточных животных (мышечные белки антин и миозин). Они выполняют также роль механической опоры. Например, высокая упругость кожи обусловлена наличием в ней коллагена.
Для белков характерна транспортная функция,. В частности, они являются транспортерами гормонов, аминокислот, липидов, саха-ров, ионов кислорода.
Белки являются источниками энергии, если происходит их распад до аминокислот. Теряя аминогруппы (дезаминируясь), белки становятся источником энергия во время, когда в клетках наступает истощение углеводных и липидных ресурсов.
Наконец, белками являются различные токсины, продуцируемые паразитами животной и растительной природы, а также змеиные яды и токсичные белки растений (рицин и другие), представляющие угрозу для животных и человека. В то же время важнейшей функцией белков является их защитная функция, поскольку иммунными антителами у животных и человека служат высокоспецифичные белки иммуноглобулины. Защитными белками можно считать также фибриноген и тромбин, которые из-за участия в свертывании крови предохраняют организм от ее потери. Кровь некоторых рыб Антарктики содержит специфический антифриз-ный белок, предотвращающий ее замерзание. Противовирусное соединение интерферон также имеет белковую природу. Наконец, некоторые белки обладают запасной пищевой функцией в семенах растений. Такие белки используются в качестве пищевых зародышами в начальный их период развития. Пищевыми белками являются альбумин (главный белок яиц птиц) и казеин (главный молочный белок).
Оценивая роль белков в жизни клеток, тканей и организмов, нельзя также не отметить, что они обладают видовой специфичностью, а это ведет к одному основополагающему заключению, сводящемуся к признанию положения «организмы делаются белками».
Нуклеиновые кислоты являются органическими соединениями, содержащими углерод, кислород, водород, азот и фосфор. Различают дезоксирибонуклеиновую и рибонуклеиновую кислоты (ДНК и РНК). Важнейшая биологическая роль нуклеиновых кислот заключается в том, что они являются хранителями генетической информации (см. главу X).
Отсылая читателя к главе X, где изложены основные сведения о нуклеиновых кислотах как генетическом материале, здесь кратко рассмотрим сведения об аденозинтрифосфорной кислоте (АТФ), которая представляет собой нуклеотид, образованный присоединением к аденозинмонофосфорной кислоте (АМФ), содержащейся в РНК, двух дополнительных молекул фосфорной кислоты (НдРО^). Другими словами, в составе АТФ содержатся аденин, рибоза и три молекулы фосфорной кислоты. АТФ синтезируется в мито-хондриях.
Выдающаяся роль АТФ определяется ее чрезвычайной важностью в обеспечении клеток энергией, которая освобождается в результате воздействия на АТФ фермента АТФ-азы, сопровождаемого в начале отщеплением одной молекулы фосфорной кислоты и образованием аденозиндифосфатной кислоты (АДФ), а затем еще двух молекул фосфорной кислоты и переходом АДФ в аденозинмо-нофосфорную кислоту (АМФ). Фосфорно-кислородные связи в АТФ (их две) называют макроэргическими, обозначая их символом Р.
Синтез АТФ происходит в митохондриях.
Углеводы — это органические соединения углерода, водорода и кислорода с общей формулой (СН2)n, где n — представляет собой число от трех до семи. Такое название этим соединениям дал в 1844 г. К. Шмидт (1822-1894). Содержание их в клетках очень значительно. Например, содержание крахмала доходит иногда до 90% сухой массы (картофель, семена злаковых).
Различают полисахариды (С6Н10О5)n, дисахариды (C12H22O11) и простые сахара — моносахариды (С6Н12О6), являющиеся малыми органическими молекулами. Две молекулы моносахарида продуцируют одну молекулу дисахарида и одну молекулу воды, тогда как из n молекул моносахарида образуется одна молекула полисахарида и (n - 1) молекул воды. Следовательно,

В зависимости от количества атомов углерода в молекуле моносахарида различают триозы (3 атома углерода), тетрозы (4 атома углерода), пентозы (5 атомов углерода) и гексозы (6 атомов углерода).
Среди триоз важное значение для животных и человека имеют такие моносахариды, как глицерин и его производные, молочная и пировиноградная кислоты.
Наиболее известной тетрозой является эритроза — промежуточный продукт фотосинтеза.
Пентозами являются рибоза и дезоксирибоза.
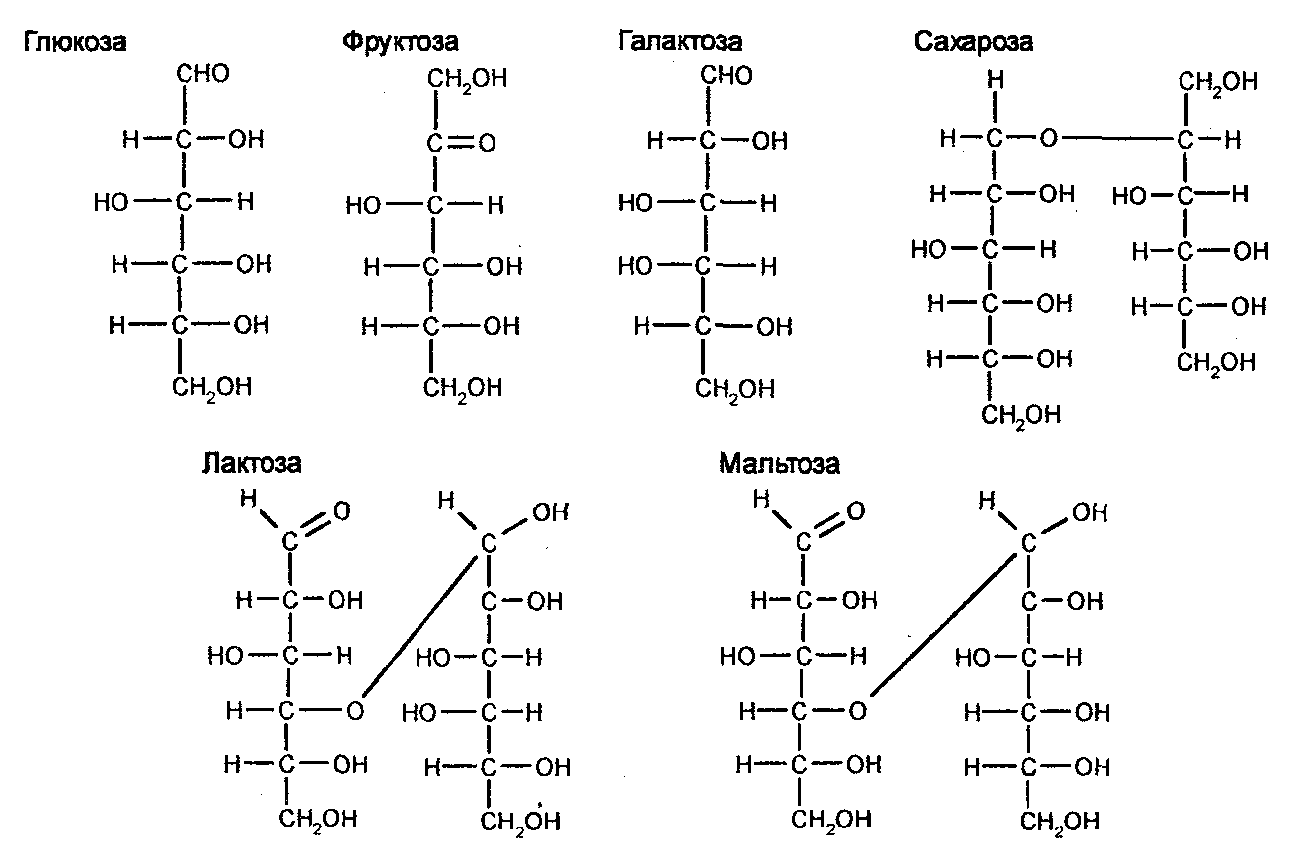
Среди гексоз важнейшее значение для организмов имеют глюкоза, являющаяся первичным источником энергии, а также фруктоза и галактоза. Глюкоза, или виноградный сахар, является основной частью ряда ди- и полисахаридов. В природных условиях в свободном состоянии она встречается в клетках практически всех растений. Что касается животных, то она также широко распространена, обнаруживаясь в структурных компонентах, в крови. В результате окисления глюкозы происходит ее распад до разных производных Сахаров, а затем и до CO2 и H2O. При этом освобождается энергия и образуется восстановительная способность, запасаемая в молекулах АТФ и НАДФ. Фруктоза (плодовый сахар) в свободном состоянии встречается в плодах растений. Особенно много ее в сахарном тростнике и сахарной свекле, во фруктах, а также меде. Галактоза встречается в составе лактозы, которой много в молоке.
Дисахариды, трисахариды и тетрасахариды часто называют оли-госахаридами, среди которых очень важными для жизни организмов являются сахароза, лактоза и мальтоза. Сахароза, известная в быту как сахар, в больших количествах содержится в сахарном тростнике и сахарной свекле. Молекулы сахарозы состоят из остатков D-глюкозы и D-фруктозы. Лактоза, или молочный сахар, в большом количестве содержится в молоке. Она состоит из глюкозы и галактозы. Мальтоза встречается в составе крахмала и гликогена. Она состоит из двух молекул глюкозы и этим определяется ее биологическая важность.
Полисахариды обладают свойствами полимеров. Будучи образованными сотнями или даже тысячами моносахаридных единиц, они являются либо линейными полимерами (целлюлоза), либо разветвленными (гликоген).
Полисахаридами, состоящими из большого количества моноса-харидов, и наиболее известными и биологически важными у растений являются целлюлоза и крахмал, которые состоят из монотонно повторяющихся остатков D-глюкозы. Являясь основным структурным элементом клеточных стенок, целлюлоза обеспечивает прочность клеток зеленых растений. Известен также полисаха-рид хитин, содержащийся в клеточных стенках грибов и в скелете членистоногих. Он обеспечивает прочность их скелета. Считают, что целлюлоза является самым распространненным углеводом среди всех углеводов, известных на Земле. Крахмал содержится в большом количестве в клубнях картофеля и семенах злаковых (особенно кукурузы и пшеницы). Он построен из двух полимеров D-глюкозы (а-амилазы и пектина). Крахмал является резервным углеводом в клетках растений. В клетках животных содержится полисахарид гликоген, который тоже состоит из очень большого количества остатков D-глюкозы. Накапливаясь в печени, мышцах и других органах, он является источником глюкозы, поступающей в кровь. Этот углевод обнаруживают также в грибах.
Полисахариды, гиалуроновая кислота и пектины создают прослойку между клетками животных и растений, соответственно. Такой полисахарид, как гепарин синтезируется клетками легких, печени и других тканей и секретируется в кровь.
Известны соединения, представляющие собой комплекс сахаров с белками. Например, небольшие олигосахаридные группы, прикрепляясь через 0-гликоидную связь к -ОН-группам остатков се-рина, треонина, оксилизина или через N-гликозидную связь к амидному азоту аспарагина формируют гликопротеиды, обладающие рядом биологических функций (ферментативных, структурных, регулирующих). Если концевое звено полисахарида ковален-тно присоединению О-гликоидной связи к сериновому остатку в белке, то образующееся сложное соединение называют пептидо-гликаном. Это соединение обладает структурной функцией.
Углеводы обладают структурной функцией, причем самым распространенным структурным углеводом является целлюлоза. Другими структурными углеводными элементами являются гликоза-миногликаны (кислые мукополисахариды) и протеогликаны.
Углеводы являются важнейшим источником энергии в организме, которая освобождается в результате окислительно-восстановительных реакций. Установлено, что окисление 1 г углевода сопровождается образованием энергии в количестве 4,2 ккал. Целлюлоза не переваривается в желудочно-кишечном тракте позвоночных из-за отсутствия гидролизующего фермента. Она переваривается лишь в организме жвачных животных (крупный и мелкий рогатый скот, верблюды, жирафы и другие). Что касается крахмала и гликогена, то в желудочно-кишечном тракте млекопитающих они легко расщепляются ферментами-амилазами. Гликоген в желудочно-кишечном тракте расщепляется до глюкозы и некоторого количества мальтозы, но в клетках животных он расщепляется гликогенфосфорилазой с образованием глюкозо-1-фосфата. Наконец, углеводы служат своеобразным питательным резервом клеток, запасаясь в них в виде гликогена в клетках животных и крахмала в клетках растений.
Липиды (от греч. lipos — жир), или жиры являются соединениями, состоящими из жирных кислот и глицерола. К этим липидам относят также жироподобные вещества (воска). Жирные кислоты — это органические кислоты. Наиболее встречаемыми жирными кислотами в жирах животных и растений являются пальмитиновая (CH3(CH2)15COOH), стеариновая (CH3(СH2)16COOH) и олеиновая (СН3-СН2)7СН-СН(СН2)7-СООН) жирные кислоты. Одна молекула глицерола и три молекулы жирной кислоты образуют одну молекулу липида и три молекулы воды. Например, стеарин образуется в результате реакции между одной молекулой глицерола и тремя молекулами стеариновой кислоты:
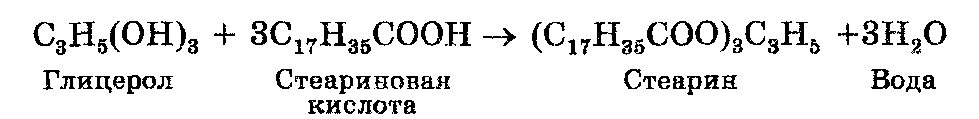
Для липидов характерно то, что они не растворимы в воде. Растворителями для них являются эфир, бензин, хлороформ и другие органические растворители.
Липиды встречаются почти во всех клетках, но в основном в небольших количествах, хотя некоторые клетки содержат эти соединения в очень больших количествах, доходящих до 90% их сухой массы. Они обнаруживаются в нервной ткани, мужских половых клетках, в семенах растений. В бараньем жире глицерол связан в основном со стеариновой кислотой, тогда как в говяжьем жире — с пальмитиновой и стеариновой кислотами. Напротив, в растительных маслах и жире лошади глицерол связан с олеиновой кислотой.
Липиды в сочетании с другими соединениями образуют более сложные соединения. Например, известны фосфолипиды (глицерол + жирная кислота + фосфатная группа), липопротеиды (комплексы липидов с белками) и другие. Названные выше липиды определяют в качестве омыляемых, т. к. нагревание их совместно со щелочами сопровождается их гидролизом с образованием мыл. Между тем известны липиды, не способные образовывать мыла. Эти липиды называют неомыляемыми или стероидами, среди которых наиболее распространенными являются стеролы (стероид-ные спирты). В тканях животных наиболее часто обнаруживается холестерол.
Липиды обладают рядом важнейших свойств в жизни клеток. Прежде всего, поскольку углеводы могут переводиться в липиды, то последние выполняют роль накопителей энергии, ибо окисление липидов сопровождается выделением энергии. Например, окисление 1 г жира сопровождается выделением энергии в количестве 9,5 ккал. При окислении образуются также углекислый газ и вода. Очень тонкий слой жира в плазматической мембране клеток выполняет защитную роль.
Очень важное значение в построении клеточных структур липиды приобрели в составе фосфолипидов, которые являются одним из основных строительных материалов мембран клеток. Важную биологическую роль в жизни клеток и организмов играют также липопротеиды. Липиды способны к запасанию в организмах в больших количествах и этим обеспечивают терморегуляцию организмов, являясь материалом для образования эндогенной воды в результате его окисления, что имеет важное значение для жизни многих животных пустынь (верблюды, мелкие млекопитающие). Являясь предшественниками в синтезе ряда гормонов, они принимают участие в регуляции важных функций организмов. Воска предохраняют кожу позвоночных от воды, у птиц они придают водоотталкивающие свойства перьям. У многих видов растений воска покрывают листья. Питаясь фитопланктоном, содержащим воска, киты и лососевые рыбы используют их в качестве главного источника липидов. Помимо соединений, рассмотренных в этом параграфе, в клетках содержатся также и другие соединения. Чтобы проявлялась каталитическая активность ферментов, для многих из них необходимо присутствие кофакторов небелковой природы, которые либо непосредственно участвуют в каталитическом процессе, либо являются промежуточными переносчиками функциональных групп от субстрата непосредственно к ферменту. Если кофакторами ферментов являются органические соединения, то их называют кофермента-ми. Предшественниками многих таких органических соединений (ко-ферментов) служат витамины, которые тоже являются органическими соединениями, присутствующими в небольших количествах в клетках растений и животных и попадающих в организм человека с пищей. К настоящему времени известно более 10 различных вята-минов, которые классифицируют на водорастворимые и жирорастворимые витамины. Водорастворимыми являются витамины B2 (тимин), B2 (рибофлавин), В6 (пиридоксин), В12 никотиновая, панто-теновая и фолиевая кислоты, биотин и витамин С (аскорбиновая кислота). Жирорастворимыми являются витамины А, Д, Е и К.
Водорастворимые витамины в качестве коферментов участвуют в катализировании многих реакций, в частности таких, как декарбоксилирование a-кетокислот (витамин В1), окислительно-восстановительных реакций (витамин В2, никотиновая кислота), реакций гидроксилирования (витамин С), карбоксилирования (витамин К), переносе ацильных (пантотеиновая кислота) и многих других реакций.
Жирорастворимые витамины выполняют самые различные функции. В частности витамин А принимает участие в формировании зрительного процесса в виде альдегида витамина А, связанного с белком опсином, витамин Д регулирует обмен кальция, витамин Е участвует в защите липидов клеточных мембран от разрушения кислородом, а витамин К является кофактором реакций карбоксилирования.
Кофакторами ферментов являются также микроэлементы. В частности, для каталитического действия многих ферментов (цитохромоксидазы, каталазы, пероксидазы) необходимы ионы железа. Для действия цитохромоксидазы и лизиноксидазы необходима медь. Ионы Zn2+ присутствуют в НАД- и НАДФ-зависимых дегидрогена-зах. Другим ферментам необходимы ионы марганца (аргиназа), никеля (уреаза) или атомы молибдена и ванадия (флавиндегидрогена-зы). Некоторые микроэлементы участвуют в регуляторных реакциях. Например, хром участвует в регуляции усвоения глюкозы клетками животных тканей, а олово необходимо для кальцификации костей. Бор и алюминий необходимы для развития растений.
Наконец, в клетках в очень небольших количествах встречаются аминокислоты в свободном состоянии (свыше 150), которые не встречаются в составе белков.
Размножение клеток
Размножение или пролиферация (от лат. proles — потомство, ferre — нести) клеток — это процесс, который приводит к росту и обновлению клеток. Данный процесс характерен как для одноклеточных, так и многоклеточных организмов.
Клетки-организмы (одноклеточные организмы) размножаются простым делением надвое (бактерии, саркодовые), множественным делением (споровики и др.) или другим путем. Поэтому у бактерий и одноклеточных животных удвоение клеток представляет собой размножение их как самостоятельных организмов, поскольку из исходной формы (организма) образуется две новые клетки, каждая из которых является организмом. Каждая дочерняя клетка (организм) получает полную генетическую информацию, несомую исходной клеткой-организмом.
Соматические клетки многоклеточных организмов размножаются путем сложного деления, которое получило название митоти-ческого деления и которое в общем виде представляет собой механизм, посредством которого одиночные клетки репродуцируют себя. Образовавшиеся в результате деления дочерние клетки подобны исходной (материнской) клетке, отличаясь от последней лишь меньшими размерами. Однако вслед за делением дочерние клетки мгновенно начинают расти и быстро достигают размеров материнской клетки.
Биологический смысл митотического деления состоит в том, что оно является ключевым событием в точной репликации всех хромосом еще до того, как произойдет деление ядра и клетки. В результате митоза дочерние клетки после деления получают хромосомы в точно таком же количестве, какое имела их родительская (материнская) клетка. Следовательно, митотическое деление есть особый способ упорядоченного деления клеток, при котором каждая из двух дочерних клеток получает хромосомы в точно таком же количестве и точно такого же строения, что и хромосомы, которые имела материнская клетка. При каждом митозе образуется копия каждой хромосомы и действует точный механизм их распределения между дочерними клетками.
В митотическом делении клетки различают две стороны — разделение исходного ядра на два дочерних ядра (равное деление хромосом), называемое кариокинезом (от греч. caryon — ядро, kinesis — движение) и представляющее собой, по существу, хромосомный цикл, и следующее затем разделение цитоплазмы с образованием двух дочерних клеток, называемое цитокинезом (от греч. cytos — клетка, kinesis — движение) и представляющее собой цитоплазматический цикл. Каждая из дочерних клеток содержит одно дочернее ядро.
Кариокинез и цитокинез протекают синхронно, причем в кариоки-незе имеет место чередование синтеза ДНК с митозом, тогда как цитокинез чередуется с ростом клеток (удвоением в числе клеточных компонентов).
Существенной особенностью митотического деления является то, что оно в значительной мере сходно у всех организмов. Совокупность процессов, происходящих в клетке от одного деления до другого, получило название митотического цикла.
Митотический цикл состоит из двух стадий — стадии покоя или интерфазы и стадии деления или митоза (от греч. rnitos — нить), обозначаемого символом м. Термины «митоз» и «кариокинез» — синонимы. Интерфаза доступна для оценки качественно и количественно, точно так же доступен для измерения и митоз. В частности, для измерения интенсивности количества митозов используют так называемый митотический индекс, под которым понимают число митозов на 1000 клеток. Данные о митотическом индексе имеют важное практическое значение, особенно в медицинской практике (в оценке интенсивности регенерации органов, действия лекарственных веществ и т. д.).
Интерфаза предшествует митозу, и функциональное содержание ее заключается в том, что в ней происходит синтез ДНК (рис. 58), причем ее длительность составляет не менее 90% в течение всего клеточного цикла. Различают три последовательных периода интерфазы, а именно: пресинтетический, синтетический и постсинтетический.
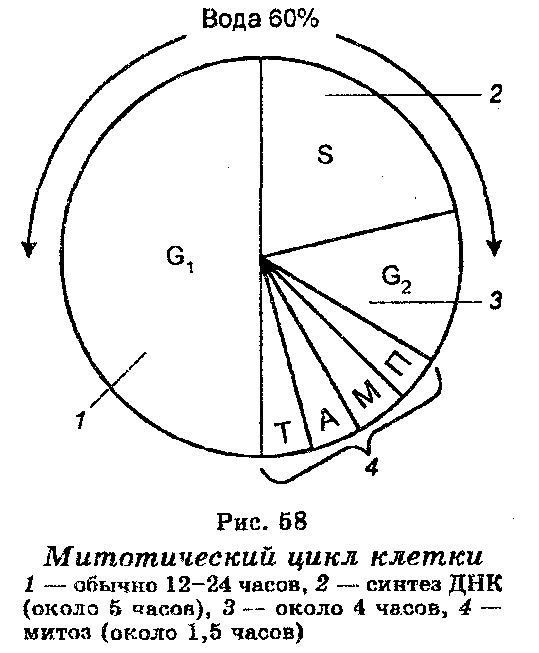 Пресинтетический период (G1), который часто называют еще первым интервалом (от англ. gap — интервал), является начальным периодом интерфазы. В этот период ДНК еще не синтезируется, однако происходит накопление РНК и белков, в том числе и белков, необходимых для синтеза ДНК. Увеличивается количество митохондрий. Обычно этот период длится 12-24 часа.
Пресинтетический период (G1), который часто называют еще первым интервалом (от англ. gap — интервал), является начальным периодом интерфазы. В этот период ДНК еще не синтезируется, однако происходит накопление РНК и белков, в том числе и белков, необходимых для синтеза ДНК. Увеличивается количество митохондрий. Обычно этот период длится 12-24 часа.
Синтетический период (S) следует за G1-периодом и характеризуется тем, что в этот период в клетке происходит синтез (репликация) ДНК, в результате чего количество ее удваивается. В этот период продолжается также синтез РНК и белков. Очень важно, что к концу этого периода каждая из хромосом удваивается и состоит уже из двух сестринских хроматид, удерживаемых центромерои. Можно сказать, что наиболее фундаментальной особенностью S-периода является репликация генов и удвоение набора генов каждой дуплицированной хромосомы. Длительность S-периода обычно составляет около 5 часов.
Постсинтетический период (G2) характеризуется остановкой синтеза ДНК и накоплением энергии. Однако продолжается синтез РНК и белков, формирующий нити веретена деления. Длительность G2-периода составляет 3—6 часов.
Митоз совершается на протяжении четырех последовательных фаз, а именно: профазы, метафазы, анафазы и телофазы (рис. 59.).
 В профазе в начале происходит конденсация и спирализация (скручивание) хромосом, в результате чего они становятся видимыми при микроскопии окрашенных препаратов. Увеличивается диаметр каждого завитка. Ядерная мембрана растворяется под действием ферментов, ядрышко исчезает. Центросома делится на две центриоли, после чего последние расходятся к полюсам клетки. Отмечается также фосфорилирование отдельных клеточных белков. Затем между полюсами начинает формироваться ахромати-новая фигура, похожая на веретено. Оно состоит из белка и РНК. К концу этой фазы ахроматиновая фигура вытягивается вдоль клетки, становясь веретеном. Структурно веретено представляет собой двухполюсную структуру, построенную из микротрубочек и различных белков. Хроматиды (сестринские хроматиды) удерживаются вместе центромерои. Длительность профазы составляет примерно 30-60 минут.
В профазе в начале происходит конденсация и спирализация (скручивание) хромосом, в результате чего они становятся видимыми при микроскопии окрашенных препаратов. Увеличивается диаметр каждого завитка. Ядерная мембрана растворяется под действием ферментов, ядрышко исчезает. Центросома делится на две центриоли, после чего последние расходятся к полюсам клетки. Отмечается также фосфорилирование отдельных клеточных белков. Затем между полюсами начинает формироваться ахромати-новая фигура, похожая на веретено. Оно состоит из белка и РНК. К концу этой фазы ахроматиновая фигура вытягивается вдоль клетки, становясь веретеном. Структурно веретено представляет собой двухполюсную структуру, построенную из микротрубочек и различных белков. Хроматиды (сестринские хроматиды) удерживаются вместе центромерои. Длительность профазы составляет примерно 30-60 минут.
В метафазе хромосомы располагаются на экваторе веретена. Они имеют вид толстых образований, плотно свернутых спиралью, что облегчает подсчет и изучение их структуры с помощью микроскопа. Будучи прикрепленными к нитям веретена центромерои, к которой прикрепляются особые белковые комплексы (кине-тофоры), связанные с отдельными микротрубочками хроматиды, пока удерживаются вместе, но плечи их уже разъединены. Длительность метафазы составляет 2—10 минут.
В анафазе наступает разделение кинетохоров, а затем и продольное разделение хромосом, в результате чего каждая сестринская хро-матида имеет собственную центромеру и становится дочерней хромосомой. Хромосомы удлиняются и двигаются к соответствующим полюсам веретена. Анафаза длится 2-3 минуты. Репликация хромосомных концов (теломер), длина которых составляет 2—20 кб., требует теломеразы.
В телофазе (от греч. telos — конец) дочерние хромосомы достигают полюсов, вытягиваются и деспирализуются. Кинетохорные трубочки исчезают. Образуется ядерная оболочка, вновь появляется ядрышко. Длительность составляет 20-30 минут.
На заключительном этапе клеточного деления происходит ци-токинез, который начинается еще в анафазе. Этот процесс заканчивается образованием в экваториальной зоне клетки перетяжки, которая разделяет делящуюся клетку на две дочерние клетки. Перетяжка обеспечивается сокращением кольца, сформированного филаментами актиновой природы.
В отличие от соматических клеток животных в клетках растений из-за ригидности их стенок вместо образования сократительного кольца формируется пластинка между будущими дочерними клетками. На каждой из сторон этой пластинки откладывается целлюлоза, после чего она становится клеточной стенкой.
Каждое клеточное деление является непрерывным процессом, поскольку ядерные и цитоплазматические фазы, вопреки различиям в содержании и по значению, координированы во времени.
Упорядоченность клеточных делений у эукариотов зависит от координации событий в клеточном цикле. У эукариот эта координация осуществляется путем регуляции трех переходных периодов в клеточном цикле, а именно: вступление в митоз, выход из митоза и прохождение через пункт, называемый «Старт», который вводит инициацию синтеза ДНК ( S-фазу) в клетке.
Продолжительность митотических циклов разных клеток различна и составляет от нескольких часов до нескольких дней. Однако она зависит от типа тканей, физиологического состояния, внешних факторов (температура, свет).
Клеточный цикл эукариотических клеток регулируется последовательной активацией циклинзависимых киназ (СДК) путем взаимодействия их с белками-циклинами. Комплекс циклин-СДК оказывается полностью активированным фосфорилированием треонинового остатка в Т-петле СДК, осуществляемым специфической СДК-активирующей киназой (САК). При этом комплекс циклин-СДК вовлекается в инициацию как митоза, так и репликации ДНК. Регуляция митоза зависит от регуляции СДК.
Существуют и другие регуляторы клеточного цикла. В частности известны регуляторы, ингибирующие СДК. Такими ингибиторами являются белки р21, р16 и р27. Они ингибируют функции киназ также путем связывания с ними.
Хромосомная ДНК в клетках организмов-эукариотов реплици-руется лишь один раз в клеточном цикле. Поэтому давно возник вопрос о механизме, ограничивающем лишь один раунд репликации ДНК в клеточном цикле. Предполагают существование так называемого лицензирующего фактора репликации (licensing factor), который позволяет репликацию. В подтверждение этого взгляда установлены лицензирующие белки MSM, которые обычно связаны с хромосомами, но с началом S-фазы освобождаются от этой связи, позволяя репликацию ДНК, а после того, как синтез ДНК завершается, вновь связывается с хромосомами.
Разные ткани характеризуются разной митотической активностью. Поэтому в зависимости от митотической активности различают стабильные, растущие и обновляющиеся ткани. Стабильные ткани — это ткани, в которых клетки не делятся, а количество клеточной ДНК постоянно. Например, клетки центральной и периферической нервной системы не делятся. В этих клетках происходят лишь возрастные изменения. Растущие ткани — это ткани, в которых клетки живут всю жизнь, но среди последних имеются такие, которые делятся посредством митоза. В результате этого наступает увеличение размеров органов. Примером растущих тканей являются ткани почек, желез внутренней секреции, скелетная и сердечная мускулатуры. Обновляющиеся ткани — это ткани, в которых многие клетки подвержены митозам, в результате чего погибающие клетки компенсируются вновь образующимися. Примерами обновляющихся тканей являются клетки желудочно-ки-шечного, дыхательного и мочеполового трактов, эпидермиса, костного мозга, семенников и др. Для митозов характерны суточные колебания, волны.
У высших организмов митотическое деление клеток обеспечивает их рост с последующим увеличением массы тела и дифференциацией клеток. По мере индивидуального развития человека количество его клеток увеличивается, достигая у взрослого человека более чем 10 клеток и оставаясь затем константным.
Как уже отмечено, митохондрии и хлоропласты способны к делению в клетках эукариотов, но контроль их деления не ясен. Установлено лишь, что в геноме клеток растений существует ген, который, возможно, принимает участие в контроле деления хло-ропластов.
Для деления клеток млекопитающих и птиц характерно то, что оно имеет определенные ограничения количества клеточных удвоений. Например, фибробласты плодов человека удваиваются лишь на протяжении 50 генераций, тогда как фибробласты от людей в возрасте 40 и 80 лет подвергаются примерно 40 и 30 удвоениям соответственно, если их культивируют в стандартных условиях. Это явление получило название старения клеток. Считают, что в организме также большинство клеток стареет, например, клетки печени живут около 18 месяцев, эритроциты — 4 месяца, в результате чего в них накапливаются липиды, кальций, пигмент «изнашивания» и они гибнут. Подсчитано, что организм взрослого человека ежедневно теряет около 1-2% своих клеток в результате их гибели. После смерти клетки в ней происходит коагуляция протоплазмы, распад митохондрий и других органелл в результате ауто-лиза (активации внутриклеточных ферментов).
Для объяснения природы старения клеток предложено несколько гипотез, в которых придается значение ошибкам биосинтетических механизмов клеток, механизмам защиты от злокачественного перерождения нормальных клеток или другим причинам. Однако ни одна из известных гипотез не является исчерпывающей в объяснении феномена старения клеток.
Установлено, что для клеток во многих случаях характерен апоптоз, под которым понимают генетическую программу, в результате которой клетки совершают суицид. Можно сказать, что апоптоз — это эволюционно сохраняемый процесс. С помощью этого процесса многоклеточные организмы освобождаются от излишних или потенциально вредных клеток. Этот феномен отличен от старения клеток. На примере нематоды Caenorhabditie elegans было выяснено, что клеточный суицид контролируется генным набором, состоящим из трех генов, контролирующих синтез белка СЕД-3, СЕД-4 и СЕД-9, регулирующих апоптоз. У млекопитающих выявлены белки-2, которые регулируют апоптозную смерть клеток. Полагают, что апоптоз имеет значение в этиологии многих наследственных болезней (болезнь Альцгеймера и Др.), аутоиммунных нарушений, сердечно-сосудистых болезней, возрастных нарушений и даже СПИДа.
Однако погибающие клетки замещаются новыми. Считают, что клеточное содержание организма человека обновляется примерно каждые семь лет. Особенно сильно замещение клеток происходит в крови за счет интенсивного образования кровяных клеток в кроветворных тканях. Применительно к другим видам клеток процесс обновления происходит с очень высокой скоростью. Например, эпителий желудка и кишечника крыс обновляется каждые 72 и 38 часов соответственно, эпителий тонкого кишечника человека — каждые 7-8 дней. Однако нервные клетки функционируют (живут) на протяжении всей жизни организмов.
Наряду с делением клеток путем митоза известен амитоз (от греч. а — не, mytosia — деление ядра), под которым понимают прямое деление ядра клетки. При амитозе сохраняется интерфазное состояние ядра, ядрышко, ядерная мембрана. Ядро клетки делится на две части без формирования веретена, в результате чего образуется двухъядерная клетка. Амитоз встречается иногда в клетках скелетной мускулатуры, кожного эпителия, соединительной ткани. Однако считают, что амитоз является аномальным механизмом в размножении клеток.
Считают, что митотический цикл у высших организмов является результатом эволюции разделительного механизма эукарио-тов. В пользу этого предположения свидетельствуют результаты сравнения разделительных механизмов бактерий, некоторых водорослей, дрожжей, простейших и млекопитающих. Это сравнение показывает, что усложнение митотического аппарата происходит по мере усложнения организации и функций организмов, принадлежащих к разным систематическим группам.
Дата добавления: 2015-12-29; просмотров: 952;