Физиологические гомеостатические механизмы.
Эта группа механизмов, включающая регуляторные функции легких, почек, желудочно-кишечного тракта, печени и костной ткани, обеспечивает нейтрализацию, связывание и выведение из организма Н+ и ОН- ионов в составе различных соединений, в зависимости от особенностей метаболизма и состояния буферных систем внутренней среды.
Легкие и кислотно-щелочное состояние.Процессы газообмена между кровью и окружающим воздухом, происходящие в легких, теснейшим образом связаны с регуляцией кислотно-щелочного состояния, поскольку внешнее дыхание — это основной путь удаления из крови летучей угольной кислоты
Количество выводимого из организма углекислого газа зависит от глубины и частоты дыхания, т.е. от альвеолярной вентиляции. Величина же альвеолярной вентиляции регулируется дыхательным центром, деятельность которого зависит или даже прямо определяется рН крови и парциальным напряжением С02. Таким образом, усиление дыхания и альвеолярной вентиляции происходит при увеличении напряжения в крови С02 и уменьшении рН. Нарастающее при этом удаление углекислоты приводит ее содержание в крови в соответствие с концентрацией бикарбоната. Следовательно, легкие выступают в роли физиологической гомеостатической системы не только выводящей избыток летучей кислоты, но и восстанавливающей емкость буферных систем, прежде всего бикарбо-натной и гемоглобиновой.
При быстром поступлении во внутреннюю среду оснований происходит уменьшение вентиляции и количество выделяемого С02 существенно падает. Концентрация углекислоты возрастает, уравнение Гендерсона-Гассельбаха восстанавливается и рН поддерживается на исходном уровне.
Участие гемоглобина в регуляции концентрации водородных ионов в крови связано с его функцией в транспорте кислорода и углекислоты. Константа диссоциации кислотных групп гемоглобина меняется в зависимости от его насыщения кислородом. При насыщении гемоглобина кислородом он становится более сильной кислотой и увеличивает отдачу в раствор ионов водорода; в том случае, когда он обедневает кислородом, наоборот - становится более слабой кислотой. Его способность связывать ионы водорода увеличивается.
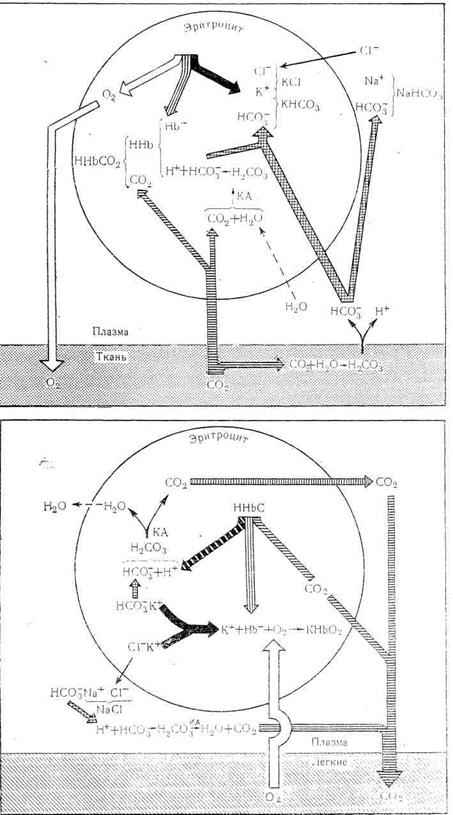
В периферических капиллярах большого круга кровообращения гемоглобин эритроцитов отдает кислород в ткани. При этом в эритроцит поступает продукт тканевого обмена-углекислый газ. Под влиянием карбоангидразы углекислый газ взаимодействует с водой, при этом образуется угольная кислота. Возникающий за счет угольной кислоты избыток водородных ионов связывается с гемоглобином, отдавшим кислород, а накапливающиеся анионы НСОз выходят из эритроцита в плазму. В обмен на эти ионы в эритроцит поступают анионы хлора, для которых мембрана эритроцита проницаема, в то время как натрий-другой составной элемент хлористого натрия, содержащегося в крови,– остается в жидкой части крови. В итоге в плазме крови повышается содержание бикарбоната натрия. Таким образом, бикарбонатная система находится в довольно тесных функциональных связях с буферной системой эритроцитов. Несмотря на то, что в кровь постоянно поступают из тканей кислые продукты метаболизма, рН крови сохраняется постоянным (рис. 2.4.1.)
Попадая в капилляры легких, буферные системы крови разгружаются от избытка кислых эквивалентов за счет выделения углекислого газа. Этому в значительной мере способствует превращение гемоглобина в оксигемоглобин. Приобретя более сильные кислотные свойства, оксигемоглобин реагирует с бикарбонатом крови с образованием Н2СО3, которая диссоциирует на СО2 + Н2О. Углекислый газ выделяется с выдыхаемым воздухом. За сутки с вдыхаемым воздухом в организм поступает около 600 л кислорода и выделяется в окружающую среду 480 л углекислого газа (примерно 21,56 моль С02).
Хотя легкие и обладают значительным влиянием на кислотно-щелочное равновесие, их эффект сказывается по истечении большего промежутка времени, чем буферных систем крови; примерно 1-3 мин требуется легким для того, чтобы сгладить наметившийся сдвиг концентрации водородных ионов в крови, тогда как буферные системы срабатывают в течение 30 с. Тем не менее, ценность легочного механизма состоит в том, что, увеличивая количество выделяемой в окружающую среду углекислоты, легкие быстро компенсируют тенденцию к ацидозу.
Важным механизмом, участвующим в регуляции постоянства концентрации водородных ионов, является почечный диурез (рис. 2.4.2).Вфизиологических условиях почки выделяют кислую мочу, рН которой колеблется от 5 до 7. Почки обеспечивают повышение или понижение концентрации бикарбонатов при изменении рН. Почки действуют медленнее, чем легкие; примерно 10-20 ч требуется почкам для того, чтобы восстановить нарушенное соотношение концентраций компонентов буферных систем.
Важнейшую роль в способности почек к выделению кислой мочи играет секреция водородных ионовклетками эпителия проксимальных и дистальных канальцев. Эта способность обусловлена наличием в клетках фермента карбангидразы,обеспечивающего образование из С02 и Н20 угольной кислоты, с последующей диссоциацией:
Н20 + С02 → Н2С03 —Н+ + НС03
Ион водорода активно секретируется с затратой энергии через апикальную мембрану клетки эпителия в просвет канальца, а из канальцевой мочи в клетку диффундирует ион натрия, обеспечивая равновесие электрических разрядов. Из клетки, уже через базальную мембрану Na+ активно с помощью Na-К-насоса удаляется в интерстиций и кровь, туда же пассивно по градиенту концентрации поступает НС03~, образуя бикарбонат натрия. Следовательно, секреция Н-ионов обеспечивает обратное всасывание бикарбоната и натрия.
|
Рис.2.4.1. Роль гемоглобина в кислотно-щелочном гомеостазе.
Причем основная часть секретируемых ионов водорода тратится именно на обеспечение реабсорбции бикарбоната. Так, секретируемый Н+-ион в просвете проксимального канальца взаимодействует с профильтровавшимся в мочу из плазмы ионом бикарбоната (в сутки около 5000 ммоль), образуя угольную кислоту. Под влиянием фермента карбангидразы, встроенного в мембрану щеточной каемки клеток эпителия канальца, угольная кислота разлагается на С02 и воду:
карбангидраза
 Н++ НСОз- → Н2С03 С02+Н20
Н++ НСОз- → Н2С03 С02+Н20
Молекулы углекислого газа легко диффундируют в клетки, где реакция катализируется в обратном направлении. Таким образом, на суточную реабсорбцию до 5000 ммоль бикарбоната расходуется эквивалентное количество секретированных ионов водорода. Выделяется же с мочой только 60 ммоль/сутки ионов водорода, преимущественно в связанном виде.
Хотя легкие и обладают значительным влиянием на кислотно-щелочное равновесие, их эффект сказывается по истечении большего промежутка времени, чем буферных систем крови; примерно 1-3 мин требуется легким для того, чтобы сгладить наметившийся сдвиг концентрации водородных ионов в крови, тогда как буферные системы срабатывают в течение 30 с. Тем не менее, ценность легочного механизма состоит в том, что, увеличивая количество выделяемой в окружающую среду углекислоты, легкие быстро компенсируют тенденцию к ацидозу.
Важным механизмом, участвующим в регуляции постоянства концентрации водородных ионов, является почечный диурез (рис. 2.4.2).Вфизиологических условиях почки выделяют кислую мочу, рН которой колеблется от 5 до 7. Почки обеспечивают повышение или понижение концентрации бикарбонатов при изменении рН. Почки действуют медленнее, чем легкие; примерно 10-20 ч требуется почкам для того, чтобы восстановить нарушенное соотношение концентраций компонентов буферных систем.
Важнейшую роль в способности почек к выделению кислой мочи играет секреция водородных ионовклетками эпителия проксимальных и дистальных канальцев. Эта способность обусловлена наличием в клетках фермента карбангидразы,обеспечивающего образование из С02 и Н20 угольной кислоты, с последующей диссоциацией:
Н20 + С02 → Н2С03 —Н+ + НС03
Ион водорода активно секретируется с затратой энергии через апикальную мембрану клетки эпителия в просвет канальца, а из канальцевой мочи в клетку диффундирует ион натрия, обеспечивая равновесие электрических разрядов. Из клетки, уже через базальную мембрану Na+ активно с помощью Na-К-насоса удаляется в интерстиций и кровь, туда же пассивно по градиенту концентрации поступает НС03~, образуя бикарбонат натрия. Следовательно, секреция Н-ионов обеспечивает обратное всасывание бикарбоната и натрия. Причем основная часть секретируемых ионов водорода тратится именно на обеспечение реабсорбции бикарбоната. Так, секретируемый Н+-ион в просвете проксимального канальца взаимодействует с профильтровавшимся в мочу из плазмы ионом бикарбоната (в сутки около 5000 ммоль), образуя угольную кислоту. Под влиянием фермента карбангидразы, встроенного в мембрану щеточной каемки клеток эпителия канальца, угольная кислота разлагается на С02 и воду:
карбангидраза
 Н++ НСОз- → Н2С03 С02+Н20
Н++ НСОз- → Н2С03 С02+Н20
Молекулы углекислого газа легко диффундируют в клетки, где реакция катализируется в обратном направлении. Таким образом, на суточную реабсорбцию до 5000 ммоль бикарбоната расходуется эквивалентное количество секретированных ионов водорода. Выделяется же с мочой только 60 ммоль/сутки ионов водорода, преимущественно в связанном виде.
Хотя легкие и обладают значительным влиянием на кислотно-щелочное равновесие, их эффект сказывается по истечении большего промежутка времени, чем буферных систем крови; примерно 1-3 мин требуется легким для того, чтобы сгладить наметившийся сдвиг концентрации водородных ионов в крови, тогда как буферные системы срабатывают в течение 30 с. Тем не менее, ценность легочного механизма состоит в том, что, увеличивая количество выделяемой в окружающую среду углекислоты, легкие быстро компенсируют тенденцию к ацидозу.
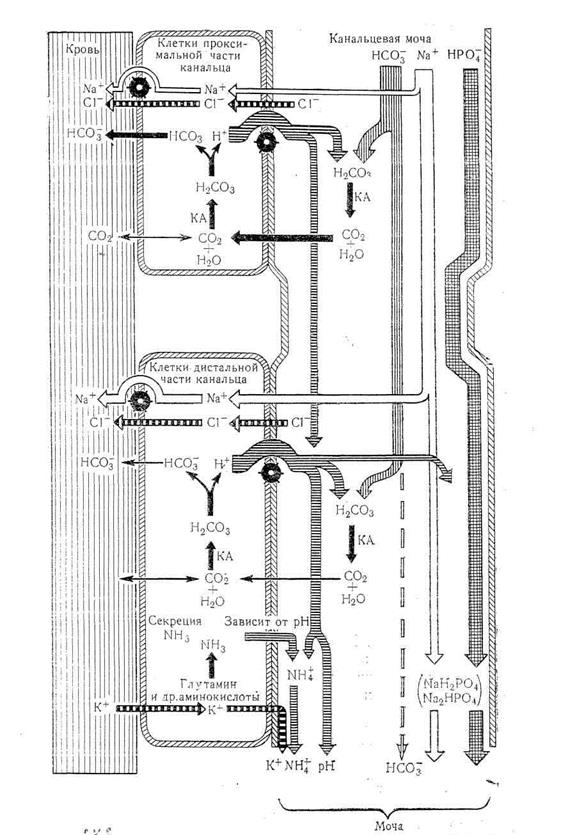
Рис. 2.4.2.Роль почек в механизмах поддержания кислотно-щелочного равновесия.
Важным механизмом, участвующим в регуляции постоянства концентрации водородных ионов, является почечный диурез (рис. 2.4.2).Вфизиологических условиях почки выделяют кислую мочу, рН которой колеблется от 5 до 7. Почки обеспечивают повышение или понижение концентрации бикарбонатов при изменении рН. Почки действуют медленнее, чем легкие; примерно 10-20 ч требуется почкам для того, чтобы восстановить нарушенное соотношение концентраций компонентов буферных систем.
Важнейшую роль в способности почек к выделению кислой мочи играет секреция водородных ионовклетками эпителия проксимальных и дистальных канальцев. Эта способность обусловлена наличием в клетках фермента карбангидразы,обеспечивающего образование из С02 и Н20 угольной кислоты, с последующей диссоциацией:
Н20 + С02 → Н2С03 —Н+ + НС03
Ион водорода активно секретируется с затратой энергии через апикальную мембрану клетки эпителия в просвет канальца, а из канальцевой мочи в клетку диффундирует ион натрия, обеспечивая равновесие электрических разрядов. Из клетки, уже через базальную мембрану Na+ активно с помощью Na-К-насоса удаляется в интерстиций и кровь, туда же пассивно по градиенту концентрации поступает НС03~, образуя бикарбонат натрия. Следовательно, секреция Н-ионов обеспечивает обратное всасывание бикарбоната и натрия. Причем основная часть секретируемых ионов водорода тратится именно на обеспечение реабсорбции бикарбоната. Так, секретируемый Н+-ион в просвете проксимального канальца взаимодействует с профильтровавшимся в мочу из плазмы ионом бикарбоната (в сутки около 5000 ммоль), образуя угольную кислоту. Под влиянием фермента карбангидразы, встроенного в мембрану щеточной каемки клеток эпителия канальца, угольная кислота разлагается на С02 и воду:
карбангидраза
 Н++ НСОз- → Н2С03 С02+Н20
Н++ НСОз- → Н2С03 С02+Н20
Молекулы углекислого газа легко диффундируют в клетки, где реакция катализируется в обратном направлении. Таким образом, на суточную реабсорбцию до 5000 ммоль бикарбоната расходуется эквивалентное количество секретированных ионов водорода. Выделяется же с мочой только 60 ммоль/сутки ионов водорода, преимущественно в связанном виде.
Наибольшая часть секретированных канальцами Н-ионов (65-75%) участвуют в процессе аммониогенеза, обеспечивающего выведение анионов сильных неорганических и органических кислот, в виде аммонийных солей. Клетки проксимальных и дистальных почечных канальцев, благодаря процессам дезаминирования и дезамидирования аминокислот, образуют аммиак. Основным источником является глутамин, который дезаминируется под влиянием фермента глутаминазы, 40% аммиака образуется из аминокислот аланина и глицина. Аммиак, благодаря высокой растворимости в липидах, легко диффундирует в просвет канальца, где присоединяет секретированные Н-ионы и образует ион аммония: NH3 + Н+ NH4+. Ионы аммония вытесняют натрий из соединений с анионами сильных кислот, натрий реабсорбируется, а анионы кислот в виде аммонийных солей выделяются с мочой.
Желудочно-кишечный тракт, печень, костная ткань и кислотно-щелочное состояние. Характер питаниячеловека и особенности пищеваренияв значительной мере сказываются на кислотно-щелочном состоянии его внутренней среды. В пищевых продуктах растительного происхождения содержится большое количество солей органических кислот (щавелевой, яблочной, лимонной, янтарной и др.), после окисления которых и выведения кислотных валентностей с углекислотой через легкие остаются и накапливаются связанные с катионами щелочные валентности (табл. 2.4.2.). Поэтому растительная пища способствует ощелачиванию внутренней среды, соответственно, основания в большем количестве выводятся с мочой, что увеличивает рН мочи и она может приобретать щелочную реакцию. Белковая пища животного происхождения способствует закислению внутренней среды, так как в составе животных белков содержатся сера и фосфор, что приводит к образованию сильных неорганических кислот. Повышенное выведение их с мочой приводит к снижению рН мочи и она приобретает резко кислую реакцию. Всасывание в желудочно-кишечном тракте кислот или щелочей, содержащихся в различных напитках, в том числе алкогольных и минеральных водах, также приводит к изменению кислотно-щелочного состояния внутренней среды.
Сравнительное небольшую роль в регуляции кислотно-щелочного состояния играют функции желудка и кишечника. Париетальные. клетки слизистой желудка секретируют соляную кислоту. Водородные ионы образуются в клетках под влиянием карбангидразы, как это описывалось выше для клеток канальцевого эпителия почек, анион хлора поступает в клетки из хлористого натрия крови путем обмена на бикарбонат. Однако при этом ощелачивания крови не происходит, т.к. НС1 желудочного сока в кишечнике всасывается обратно в кровь. Значительное количество бикарбонатов поступает в кишечник в составе сока поджелудочной железы и кишечного сока. При этом ионы IV всасываются в кровь. Обратное всасывание претерпевают и бикарбонатные ионы, поэтому сдвигов кислотно-щелочного состояния крови не происходит. При кислой диете увеличивается выделение двухвалентных катионов Са2+ и Mg2+, a при щелочной — и одновалентных катионов. Хотя значение желудочно-кишечного тракта в физиологической регуляции кислотно-щелочного состояния очень невелико, тем не менее при изменениях, а тем более нарушениях функций желудка и кишечника неизбежно происходят сдвиги и кислотно-щелочного состояния. Стойкое и резкое повышение кислотности желудочного сока может вести к увеличение щелочного резерва крови. Частая рвота, особенно при непроходимости кишечника, вызывая потерю Н+-ионов и хлоридов вызывает резкие сдвиги в крови, требующие обязательной врачебной коррекции.
В печенипроисходит утилизация молочной кислоты с образованием гликогена, здесь разрушаются кетоновые тела (ацетоуксусная и бета-оксимасляная кислоты), происходят разнообразные метаболические превращения органических кислот и оснований. Образуя желчь с разной степенью щелочной реакции печень способствует удалению из внутренней среды разных количеств оснований.
В нейтрализации кислот существенную роль играет костная ткань, поскольку в ней связано огромное количество катионов и фосфата.
Таблица2.4.2.
 Продукты, окисляющие или ощелачивающие организм (no H. 0. Уокеру и Р. Д. Поупу)
Продукты, окисляющие или ощелачивающие организм (no H. 0. Уокеру и Р. Д. Поупу)
| Продукты | Окисление | Ощелачивание |
| Бобы свежие | ||
| Бобы сушеные | — | |
| Бобы запеченные | ||
| Ячневая крупа | — | |
| Ячмень | __. | |
| Крахмал | _ | |
| Овсяная крупа | __ | |
| Мамалыга и кукуруза | ||
| хлопья | — | |
| Хлеб черный | -— | |
| Хлеб белый | — | |
| Мука белая | — | |
| Молоко цельное | — | |
| Сыворотка молочная | - | |
| Земляные орехи | — | |
| Миндаль | — | |
| Сыр твердый | __ | |
| Сыр мягкий | — | |
| Сливки | — | |
| Яйца (в целом) | — | |
| Яйца (белок) | — | |
| Говядина | — | |
| Телятина | — | |
| Печень говяжья | —- | |
| Цыплята | — | |
| Дичь | О … 0000 | — |
| Гаранина варен. | — | |
| Баранина тушен. | — | |
| Ветчина постная свеж. | —- | |
| Бекон ж при. | — | |
| Бекон топь | — | |
| Свинина не ж и р. | ||
| Сало свиное | ._„. | |
| Рыба (всякая) | 00-000 | — |
| Палтус | — | |
| Раки | — | |
| Устрицы | -- | |
| Мидии | — | |
| Абрикосы свежие | ||
| Абрикосы сушеные | ||
| Яблоки свежие | ||
| Яблоки сушеные | ||
| Бананы спелые | ||
| Бананы зеленые | ||
| Виноград | ||
| Виноградный сок натур. | ||
| Виноградн. сок подслащ. | ||
| Сливы сушеные | ||
| Сливы маринов. | ||
| Персики | ||
| Вишни | ||
| Сок лимонн. натур. | ||
| Сок лимонн. подслащ, | ||
| Сок апельс, натур. | ||
| Арбузы | ||
| Дыни | ||
| Чернослив | ||
| Изюм | ||
| Финики | ||
| Инжир сушеный | ||
| Смородина | ||
| Клюква | ||
| Ягоды (всякие) | ||
| Фрукты (почти все) | ||
| Фрукты, вареные с сахар | 0…000 | |
| Капуста | ||
| Цветная капуста | ||
| Сельдерей | ||
| Огурцы свежие | ||
| Одуванчик (зелень) | ||
| Латук | ||
| Лук | ||
| Пастернак | ||
| Зеленый горошек | ||
| Горох сухой | ||
| Редис | ||
| Перцы | ||
| Помидоры свежие | ||
| Свекла свежая | ||
| Морковь | ||
| Картофель с кожурой |
Условные обозначения: 0 - слабое окисление или ощелачивание; 00 — среднее; 000 — сильное; 0000 - очень сильное.
Основные физиологические показатели кислотно-щелочного равновесия.
Кислотно-щелочное состояние внутренней среды характеризуют следующие показатели: актуальный рН; парциальное напряжение углекислоты; актуальный бикарбонат крови; стандартный бикарбонат крови; буферные основания крови; избыток или дефицит буферных оснований крови.
Таблица 2.4.3.
Лабораторные показатели КЩР
| ПОКАЗАТЕЛИ | ||
| Международная символика | Отечественная символика | Норма |
| рН крови | рН | 7,4 |
| рСО2 крови | рСО2 | 40 мм рт.ст. |
| концентрация Нb | концентрация Нb | 160 г/л |
| ВВ (Bufer base) | БО — буферные основания, щелочной резерв | 48-49 ммоль/л* |
| BD (Base deficite) | ДО — дефицит буферных оснований | 0-2 ммоль/л** |
| BE (Base excess) | ИО — избыток буферных оснований | 0-2 ммоль/л** |
| SB (Standart bicarbonate) | СБ — стандартные бикарбонаты | при рН 7, 4 и рС02 = 40 мм рт.ст. СБ = 24 ммоль/л*** |
| AB (Actual bicarbonate) | АБ — актуальные, или истинные, бикарбонаты | без приведения к стандартным условиям, 23 ммоль/л |
Пояснения к таблице:
БО - буферные основания включают следующие показатели:
бикарбонатный буфер - 24 ммоль/л; протеиновый буфер - 17 ммоль/л (при условии, что концентрация белка в крови 70 г/л), гемоглобиновый буфер - 6,7ммоль/л (при условии, что концентрация гемоглобина - 160 г/л), фосфатный - 2 ммоль/л: общая сумма БО - 48-49 ммоль/л.
** При анализе показателей КЩР рассчитывают разницу между нормальными величинами БО и реальными (актуальными). Разница между ними указывает на наличие дефицита оснований ДО, или избытка оснований ИО соответственно. ДО измеряется в ммоль/л и означает количество оснований (в ммоль/л), необходимое для приведения рН к 7,4. ИО - избыток оснований - означает количество кислоты, необходимой для приведения рН к 7,4 (в ммоль/л).
Дата добавления: 2015-12-16; просмотров: 1026;