Типы нуклеиновых кислот и их функции
Из двух типов нуклеиновых кислот — ДНК и РНК — дезоксирибонуклеиновая кислота выполняет роль вещества, в котором закодирована вся основная наследственная информация клетки, и которое способно к самовоспроизведению, а рибонуклеиновые кислоты выполняют роль посредников между ДНК и белком. Такие функции нуклеиновых кислот тесно связаны с обенностями их индивидуальной структуры.
ДНК и РНК — это полимерные макромолекулы, мономерами которых служат нуклеотиды. Каждый нуклеотид сформирован из трех частей — моносахарида, остатка фосфорной кислоты и азотистого основания. Азотистое основание соединено с сахаром b-N-гликозидной связью (рис. 1.1).
Сахар, входящий в состав нуклеотида (пентоза), может присутствовать в одной из двух форм: b-D-рибозы и b-D-2-дезоксирибозы. Различие между ними состоит в том, что гидроксил рибозы при 2’-углеродном атоме пентозы замещен в дезоксирибозе на атом водорода. Нуклеотиды, содержащие рибозу, называются рибонуклеотидами и являются мономерами РНК, а нуклеотиды, содержащие дезоксирибозу, носят название дезоксирибонуклеотиды и формируют ДНК.
Азотистые основания являются производными одного из двух соединений — пуринаили пиримидина. В нуклеиновых кислотах преобладают два пуриновых основания — аденин (А) и гуанин (G) и три пиримидиновых — цитозин (С), тимин (Т) и урацил (U). В рибонуклеотидах и соответственно в РНК присутствуют основания А, G, С, U, а в дезоксирибонуклеотидах и в ДНК — А, G, С, Т.

Рис. 1.1. Структура нуклеозида и нуклеотида: цифрами обозначено по-
ложение атомов в остатке пентозы
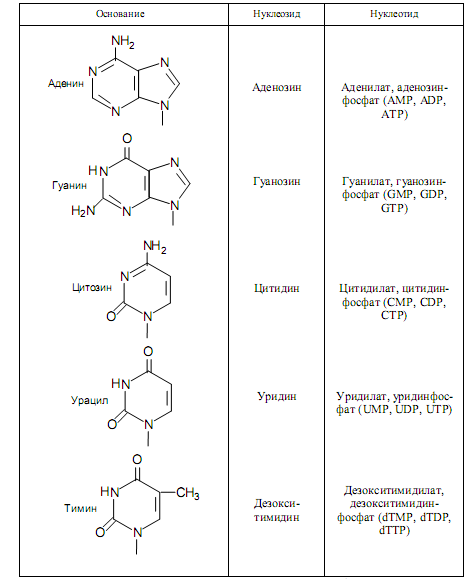
Номенклатура нуклеозидов и нуклеотидов широко используется в биохимии и молекулярной биологии и представлена в табл. 1.1.
Таблица 1.1. Номенклатура нуклеотидов и нуклеозидов
Длинные полинуклеотидные цепочки ДНК и РНК образуются при соединении нуклеотидов между собой с помощью фосфодиэфирных мостиков. Каждый фосфат соединяет гидроксил при 3’-углеродном атоме пентозы одного нуклеотида с ОН-группой при 5’-углеродном атоме пентозы соседнего нуклеотида (рис. 1.2).
При кислотном гидролизе нуклеиновых кислот образуются отдельные компоненты нуклеотидов, а при ферментативном гидролизе с помощью нуклеаз расщепляются определенные связи в составе фосфодиэфирного мостика и при этом обнажаются 3’- и 5’-концы молекулы (рис. 1.2).

Это дает основание считать цепочку нуклеиновой кислоты полярной, и появляется возможность определять направление чтения последовательности нуклеотидов в ней. Следует отметить, что большинство ферментов, участвующих в синтезе и гидролизе нуклеиновых кислот, работают в направлении от 5’- к 3’-концу (5’ → 3’) цепочки нуклеиновой кислоты. Согласно принятому соглашению, последовательность нуклеотидов в цепочках нуклеиновых кислот тоже читается в направлении 5’ → 3’ (рис. 1.2).
Особенности строения ДНК. Согласно трехмерной модели, предложенной Уотсоном и Криком в 1953 г., молекула ДНК состоит из двух полинуклеотидных цепей, которые образуют правую спираль относительно одной и той же оси. Направление цепей в молекуле взаимно противоположное, она имеет почти постоянный диаметр и другие параметры, которые не зависят от нуклеотидного состава, в отличие от белков, у которых последовательность аминокислотных остатков определяет вторичную и третичную структуру молекулы.
Сахарофосфатный остов располагается по периферии спирали, а азотистые основания находятся внутри, и их плоскости перпендикулярны оси спирали. Между основаниями, расположенными друг напротив друга в противоположных цепях, формируются специфические водородные связи: аденин всегда связывается с тимином, а гуанин с цитозином. Причем в АТ-паре основания соединены двумя водородными связями: одна из них образуется между амино- и кетогруппами, а другая — между двумя атомами азота пурина и пиримидина соответственно. В GС-паре имеется три водородные связи: две из них образуются между амино- и кето-группами соответствующих оснований, а третья — между атомом азота пиримидина и водородом (заместителем у атома азота) пурина.
Таким образом, более объемные пурины всегда спариваются с пиримидинами, имеющими меньшие размеры. Это приводит к тому, что расстояния между С1’-атомами дезоксирибозы в двух цепях оказываются одинаковыми для АТ- и GС-пар и равными 1,085 нм. Два указанных типа пар нуклеотидов, АТ и GС, называют комплементарными парами. Образование пар между двумя пуринами, двумя пиримидинами или некомплементарными основаниями (А+С или G+Т) стерически затруднено, поскольку при этом не могут образовываться подходящие водородные связи и, следовательно, нарушается геометрия спирали.
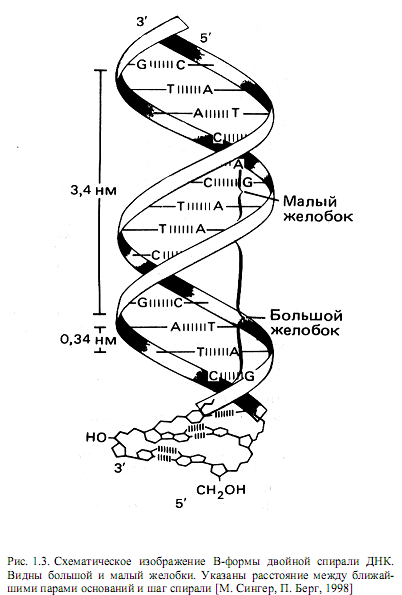
Геометрия двойной спирали такова, что соседние нуклеотиды в цепи находятся друг от друга на расстоянии 0,34 нм. На один виток спирали приходится 10 пар нуклеотидов, и шаг спирали равен 3,4 нм (10 * 0,34 нм). Диаметр двойной спирали равен примерно 2,0 нм. В связи с тем, что сахарофосфатный остов расположен дальше от оси спирали, чем азотистые основания, в двойной спирали имеются желобки —большой и малый (рис. 1.3).
Молекула ДНК способна принимать различные конформации. Обнаружены А-, В- и Z-формы. В-ДНК — это обычная форма, в которой ДНК находится в клетке, в ней плоскости колец оснований перпендикулярны оси двойной спирали. В А-форме ДНК плоскости пар оснований повернуты примерно на 20° от нормали к оси правой двойной спирали. Z-форма ДНК — это левая спираль с 12 парами нуклеотидов на виток. Биологические функции А- и Z- форм ДНК до конца не выяснены.
Стабильность двойной спирали обусловлена водородными связями между комплементарными нуклеотидами в антипараллельных цепях, стэкинг-взаимодействием (межплоскостные вандерваальсовы контакты между атомами и перекрывание p-орбиталей атомов контактирующих оснований), а также гидрофобными взаимодействиями. Последние выражаются в том, что неполярные азотистые основания обращены внутрь спирали и защищены от непосредственного контакта с полярным растворителем, и наоборот, заряженные сахарафосфатные группы обращены наружу и контактируют с растворителем.
Поскольку две цепи ДНК связаны между собой только нековалентными связями, молекула ДНК легко распадается на отдельные цепочки при нагревании или в щелочных растворах (денатурация). Однако при медленном охлаждении (отжиг) цепи способны вновь ассоциировать, и между комплементарными основаниями восстанавливаются водородные связи (ренатурация). Эти свойства ДНК имеют большое значение для методологии генетической инженерии (глава 20).
Размер молекул ДНК выражают в числе пар нуклеотидов, при этом за единицу принимается тысяча пар нуклеотидов (т. п. н.) или 1 килобаза (кб). Молекулярная масса одной т. п. н. В-формы ДНК составляет ~ 6,6*105 Да, а ее длина составляет 340 нм. Полный геном Е.coli (~ 4*106 п. н.) представлен одной кольцевой молекулой ДНК (нуклеоид) и имеет длину 1,4 мм.
Особенности строения и функции РНК. Молекулы РНК представляют собой полинуклеотиды, состоящие из одной цепи, включающей 70— 10000 нуклеотидов (иногда и больше), представленные следующими типами: мРНК (матричная или информационная), тРНК (транспортная), рРНК (рибосомная) и только в клетках эукариот — гяРНК (гетерогенная ядерная), а также мяРНК (малые ядерные). Перечисленные виды РНК выполняют специфические функции, кроме того, в некоторых вирусных частицах РНК является носителем генетической информации.
Матричная РНК является транскриптом определенного фрагмента смысловой цепи ДНК и синтезируется в ходе транскрипции. мРНК — это программа (матрица), по которой строится полипептидная молекула. Каждые три последовательно расположенных нуклеотида в мРНК выполняют функцию кодона, определяя положение соответствующей аминокислоты в пептиде. Таким образом, мРНК служит посредником между ДНК и белком.
Транспортная РНК также участвует в процессе синтеза белка. Ее функция состоит в доставке аминокислот к месту синтеза и определении положения аминокислоты в пептиде. Для этого в составе тРНК имеется специфический триплет нуклеотидов, носящий название «антикодон», и вся молекула характеризуется уникальным строением. Структурное представление о молекуле тРНК носит название «клеверный лист» (рис. 1.4).
Молекула тРНК — короткая и состоит из 74—90 нуклеотидов. Как и любая цепь нуклеиновой кислоты, она имеет 2 конца: фосфорилированный 5’-конец и 3’-конец, на котором всегда присутствуют 3 нуклеотида —ССА и концевая 3’ОН-группа. К 3’-концу тРНК прикрепляется аминокислота, и он называется акцепторным. В составе тРНК обнаружено несколько необычным образом модифицированных нуклеотидов, не встречающихся в других нуклеиновых кислотах.

Несмотря на то, что молекула тРНК одноцепочечная, в ней присутствуют отдельные дуплексные участки, формирующие т. н. стебли или ветви, где между асимметричными участками цепи образуются Уотсон—Криковские пары (рис. 1.4). Все известные тРНК формируют «клеверный лист» с четырьмя стеблями (акцепторным, D, антикодоновым и Т). Стебли имеют форму правой двойной спирали, известной как А-форма ДНК. Петли тРНК представляют собой одноцепочечные участки. Некоторые тРНК имеют дополнительные петли и/или стебли (например, вариабельная петля дрожжевой фенилаланиновой тРНК).
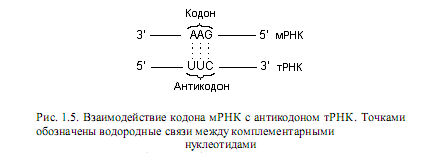
Узнавание молекулой тРНК соответствующего сайта в мРНК осуществляется с помощью антикодона, расположенного в антикодоновой петле рис. 1.4). При этом образуются водородные связи между основаниями кодона и антикодона, при условии, что формирующие их последовательности комплементарны, а полинуклеотидные цепи антипараллельны (рис. 1.5).
Молекулы разных тРНК отличаются друг от друга последовательностью нуклеотидов, однако их третичная структура очень сходна. Молекула имеет такой характер укладки, что напоминает по форме букву Г. Акцепторный и Т-стебли уложены в пространстве особым образом и образуют одну непрерывную спираль — «перекладину» буквы Г; антикодоновый и D-стебли образуют «ножку». Правильная укладка молекул тРНК в пространстве имеет большое значение для их функционирования.
В количественном отношении в клетке преобладает рибосомная РНК, однако ее разнообразие по сравнению с другими типами РНК —наименьшее: на долю рРНК приходится до 80 % массы клеточных РНК, и она представлена тремя—четырьмя видами. В то же время, масса почти 100 видов тРНК составляет около 15 %, а доля нескольких тысяч различных мРНК — менее 5 % массы клеточной РНК.
В клетках E.coli обнаружено 3 типа рРНК: 5 S, 16 S и 23 S, а в эукариотических клетках функционируют 18 S-, 5,8 S-, 28 S- и 5 S-рРНК. Эти виды рРНК входят в состав рибосом и составляют примерно 65 % их массы. В составе рибосом рРНК плотно упакованы, способны складываться с образованием стеблей со спаренными основаниями, подобными таковым в тРНК. Считается, что рРНК принимают участие в связывании рибосомы с тРНК. Показано, в частности, что 5 S-рРНК взаимодействует с Т-плечом тРНК.
Кроме перечисленных типов РНК, у эукариот в ядрах обнаружены гетерогенные ядерные РНК и малые ядерные РНК. На долю гяРНК приходится менее 2 % от общего количества клеточной РНК. Эти молекулы способны к быстрым превращениям — для большинства из них время полужизни не превышает 10 мин. Одной из немногих выявленных функций гяРНК является ее роль в качестве предшественника мРНК. мяРНК
ассоциированы с рядом белков и формируют так называемые малые ядерные рибонуклеопротеидные частицы (мяРНП), осуществляющие сплайсинг РНК (глава 3).
Дата добавления: 2015-09-29; просмотров: 4654;