Структура и функции гена. Организация генов в хромосомах
Термин «ген» ввел датский ученый В. Иоганнсен в 1909 г., когда еще не была известна его материальная природа. Пониманию структуры и функций гена способствовали результаты экспериментов американских исследователей Дж. Бидла и Э. Тейтума, которые изучали биохимическую роль различных генов у гриба Neurospora crassa. Было установлено однозначное соответствие между появлением генетической мутации, индуцированной рентгеновским излучением, и исчезновением определенного фермента, необходимого для данной биохимической стадии метаболизма. Исходя из этого, Бидл и Тейтум сформулировали гипотезу «один ген — один фермент», которая означала, что каждый ген направляет синтез одного фермента. В настоящее время эта гипотеза претерпела лишь одно изменение, связанное с тем, что структура некоторых белков, включающих более чем одну полипептидную цепь, кодируется несколькими генами. При этом последовательность аминокислот в каждой полипептидной цепи кодируется отдельным геном, цепи синтезируются отдельно и лишь затем соединяются в готовый продукт. Чаще гены, контролирующие синтез двух или нескольких полипептидных цепей, располагаются рядом на хромосоме, но не всегда. Так, например, гены, определяющие структуру a- и b-цепей гемоглобина, не сцеплены между собой. Таким образом, современная трактовка постулата Бидла и Тейтума звучит «один ген — одна полипептидная цепь». Это открытие позволило вплотную подойти к расшифровке механизмов реализации генетической информации.
В настоящее время ген понимают как структурную единицу наследственной информации, далее неделимую в функциональном отношении. Ген — это участок молекулы ДНК (реже, только у некоторых вирусов —РНК), кодирующий структуру одной макромолекулы: полипептида, тРНК или рРНК. В структуре генов прокариот, эукариот и вирусов, а также в организации этих генов в хромосомах много общего, однако есть и существенные различия.
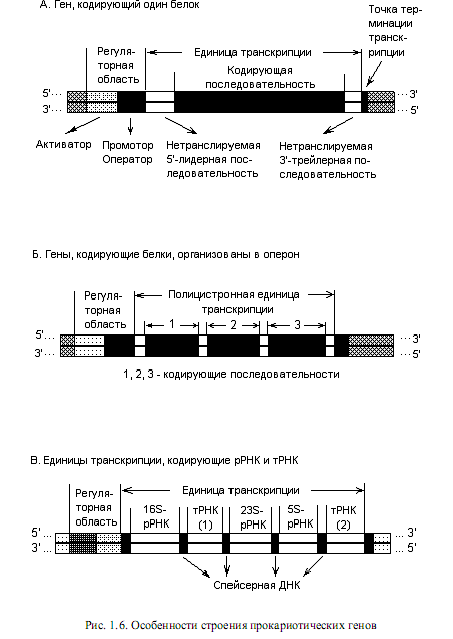
Нуклеоид прокариот содержит примерно 2—3 тыс. не перекрывающихся генов. Среди них выделяют независимые геныи гены, организованные в группы. Независимые гены называются так потому, что мРНК, считанная с такого гена, всегда моноцистронная (под цистроном понимают последовательность нуклеотидов, кодирующую единую полипептидную цепь или стабильную РНК). В свою очередь, независимые гены у прокариот могут содержать регуляторные области (рис. 1.6, А), и в таком случае их транскрипцияподвержена регуляции; а могут и не содержать таковых. В последнем случае они носят название конститутивных генов, поскольку их транскрипция осуществляется непрерывно (конститутивно), независимо от ситуации в клетке. Конститутивные гены кодируют структуру конститутивных белков.
В большинстве случаев, однако, единицы транскрипции прокариот являются полицистроннымии содержат последовательности, кодирующие не один, а несколько типов белков или РНК (рис. 1.6, Б и В). Как правило, транскрипция кодирующих последовательностей в полицистронной единице осуществляется согласованно, с участием общих 5’- и 3’-регуляторных элементов. При этом последовательности, кодирующие один или несколько полипептидов, транскрибируются с образованием зрелой мРНК, которая не претерпевает событий модификации перед трансляцией. Наоборот, последовательности, кодирующие разные типы РНК, специфически расщепляются в ходе посттранскрипционного процессинга с образованием зрелых стабильных РНК-продуктов.
Таким образом, современное представление о прокариотическом гене распространяется на следующие элементы:
1) единицы транскрипции, включающие последовательности, кодирующие зрелую РНК, либо полипептид, 5’-лидерную и 3’-трейлерную последовательности, а также спейсерную ДНК;
2) 5’-последовательности, необходимые для начала правильной транскрипции (промотор) и 3’-последовательности, нужные для правильного окончания транскрипции (терминатор);
3) последовательности, регулирующие частоту инициации транскрипции.
На рис. 1.6 показаны все перечисленные элементы, входящие в состав разных прокариотических генов. Единица транскрипции представляет собой участок ДНК между сайтами, в которых начинается и заканчивается транскрипция. Для белок-кодирующих генов характерно наличие в составе транскрипционной единицы определенного количества нуклеотидов, которые предшествуют белок-кодирующей последовательности (5’-лидер) или следующих за ней (3’-трейлер). Эти элементы присутствуют в зрелых мРНК, известно участие 5’-лидерной последовательности в процессе регуляции транскрипции.
Спейсерная ДНК представляет собой промежуточные последовательности, разделяющие кодирующие области, и она удаляется в ходе процессинга первичных транскриптов. Последовательности, необходимые для правильного начала транскрипции, представляют собой, прежде всего, промотор, с которым связывается РНК-полимераза, и участки, влияющие на скорость инициации транскрипции (оператор, активатор). Нуклеотидные последовательности, ответственные за терминацию транскрипции, располагаются на 3’-конце гена.
В нуклеоиде гены почти непрерывно следуют один за другим по всей длине ДНК, а иногда (в очень редких случаях) даже перекрываются. Значительная часть прокариотических генов объединена в группы по функциональному признаку. Например, гены путей биосинтеза аминокислот, путей катаболизма углеводов у прокариот часто объединяются в опероны. В этом случае их экспрессия осуществляется согласованно.
Число генов в геномах эукариот обычно на порядок больше, чем у прокариот. Например, в геноме человека по разным оценкам насчитывается 40— 60*103 генов. Организация в эукариотических хромосомах и сама структура генов характеризуются некоторыми отличительными особенностями. В первую очередь, у эукариот в процессе транскрипции принимает участие не один тип РНК-полимеразы, как это имеет место в прокариотических
клетках, а несколько разных ферментов. Поэтому сами единицы транскрипции и их регуляторные последовательности отличаются большей сложностью и разнообразием структурных элементов. Во-вторых, в составе эукариотических генов изобилуют мозаичные единицы транскрипции, в которых чередуются кодирующие (экзоны) и некодирующие (интроны) последовательности. Интроны чаще всего встречаются в генах, кодирующих полипептиды и тРНК, и реже в рРНК-генах. Размеры, число и местоположение интронов у разных генов различны. В целом общая длина последовательностей интронов превышает суммарную длину экзонов в 2— 10 раз и больше. Интроны вырезаются из состава мРНК в процессе сплайсинга. Третья особенность эукариотических генов состоит в том, что все белок-кодирующие мРНК у них – моноцистронные, не сгруппированные в опероны. Гены 5 S- рРНК располагаются в хромосомах эукариот тандемно (следуя один за другим в количестве нескольких копий), но каждый ген транскрибируется со своего собственного промотора с образованием РНК, имеющей только одну последовательность 5 S-рРНК на молекулу. Напротив, остальные типы рРНК образуют кластеры (группы тесно расположенных генов с общим промотором) и транскрибируются в виде полицистронной молекулы РНК, из которой в ходе посттранскрипционного процессинга образуются зрелые молекулы 18 S-, 5,8 S- и 28 S-рРНК. Количество генов в геномах разных эукариот сильно отличается, приближаясь к 105 .
Количество генов в вирусных геномах самое маленькое — обычно до десяти. Их особенностью является способность к перекрыванию в результате использования нескольких рамок считывания генетического кода. При таком способе записи наследственной информации увеличивается емкость генетического материала, что необходимо вирусам из-за ограниченных размеров капсидов, в которые может поместиться строго определенное количество нуклеиновых кислот.
Дата добавления: 2015-09-29; просмотров: 2866;