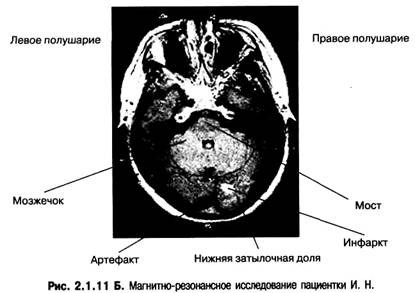
Горизонтальный срез через нижнюю затылочную долю, мозжечок и мост.
На Т2-образах заметен небольшой инфаркт в нижней затылочной доле левого полушария. Отсутствие зрительной агнозии незамаскированных объектов в данном случае может быть связано с небольшим размером инфаркта, который вызвал только агнозию скрытых объектов неконвенционального типа.
Серьезные расстройства распознания скрытых объектов у затылочных пациентов можно объяснить ролью нарушений промежуточных стадий переработки конвенциональной информации, а именно, стадий завершения. Вопросы локализации данных поражений в затылочной доле можно обсуждать с привлечением данных о записи активности одиночных нейронов (single cell recording) и удалении конкретных областей мозга у обезьян.
В определенных нейронах области V2 (18 поле Бродмана в затылочной коре, см. обзор в von der Heydt, 1995) была зафиксирована активность в ответ на «реальные» и иллюзорные, «субъективные» контуры, заданные углами и линиями. Поражения области V2 также могут быть связаны с расстройствами процесса завершения, который необходим для распознания простых свойств, замаскированных шумом. Мериган и др. показали, что изолированное поражение области V2 у обезьян приводит к нарушениям различения трех коротких вертикальных и горизонтальных линий, замаскированных перпендикулярными по отношению к ним линиями (Merigan et al., 1993). Также наблюдалось увеличение порогов различения пяти коллинеарных рядов точек, замаскированных случайно расположенными точками. Аналогичные нарушения распознания объектов, замаскированных «конкурирующим» стимулом, обнаружены у животных после удаления области V4 (Schilling, 1993).
Ограниченные поражения области V4 у приматов нарушали различение форм и шаблонов при сохранности порогов ахроматической яркости (Heywood, Cowey, 1987; Heywood et all, 1992). Они также приводили к серьезным расстройствам различения формы, требующего опоры на многочисленные признаки (Merigan, Pham, 1998), хотя нарушения различения элементарных зрительных стимулов были весьма умеренными (Schiller, 1993; Merigan, 1993; Merigan, Pham, 1998; см. обзор в Pollen, 1999).
Эти данные указывают на возможность того, что поражения областей V2 и V4 у пациентов со зрительной агнозией объектов могут приводить к нарушениям процесса завершения, необходимого для различения контуров и распознания объектов.
Поражения височной доли.Переработка конвенциональной (обычной) информации. Ассоциативная агнозия. Структуры височной доли могут участвовать в процессе сравнения объекта с моделью памяти в ходе распознания знакомых объектов. Поражения этих областей височной доли, особенно задних участков нижневисочной, веретенообразной и перигиппокампальной извилин могут участвовать в развитии ассоциативных типов зрительной агнозии объектов. Такая возможность подтверждается частым распространением затылочных поражений на нижневисочную область в большинстве случаев ассоциативной агнозии. Поражения височной доли были обнаружены в 11 из 12 случаев ассоциативной агнозии и только в 3 из 9 случаев апперцептивной агнозии. Это, возможно, указывает на особую роль поражений задней нижневисочной области, сопровождающихся затылочными поражениями, в развитии ассоциативной агнозии. Этот тип агнозии характеризуется расстройствами сравнения описания объекта с его значениями. При апперцептивной агнозии процесс кодировки и описания объекта нарушает-
ся, и эти нарушения могут быть связаны главным образом с поражениями затылочной доли.
Переработка неконвенциональной (необычной) информации.Особое внимание следует уделить роли поражений височной доли в развитии неконвенциональных типов зрительной предметной агнозии. Существуют значительные расхождения между клинико-анатомическими описаниями случаев зрительной предметной агнозии и результатами нейропсихологических исследований, прежде всего исследований, основанных на записи потенциалов одиночных нейронов, вызванных зрительным стимулом.
Клинико-анатомические данные указывают на главенствующую роль в развитии зрительной агнозии конвенциональных и неконвенциональных объектов поражений затылочной доли, местами затрагивающих задненижние участки височной доли. В то же время, исследования одиночных нейронов предоставляют нам достаточные свидетельства того, что различные нейроны нижневисочной (НВ) области реагируют на конкретные объекты, лица, части лица, руки независимо от их ориентации, размера, цвета и особенностей освещения (см. обзор в Young, 1995), а также от изменения позиции наблюдателя (Logothetis, 1995). Некоторые авторы подчеркивают, что «нейронная сеть НВ у приматов, а также пространственно-временная связь множества классов нервных клеток является представительством объекта» (Grusser, Landis, 1991, с. 126). Другие авторы восторженно объявляют, что «передняя нижневисочная область коры у обезьян является последней стадией зрительного коркового потока, отвечающего за распознание объектов» (Fujita et al., 1992, с. 343). Эти расхождения можно частично объяснить различиями в переработке конвенциональной и неконвенциональной зрительной информации.
Нарушения зрительной функции были обнаружены у обезьян после билатеральной резекции нижневисочной области, средней и нижней височных извилин (Chow, 1951; Mishkin, Pribram 1954; Wilson, 1957; Mishkin, 1966). После ампутации верхней височной извилины, височного полюса, миндалины, гиппокампальной или заднете-менной области, фронтальной коры, подушки таламуса, дорсомедиального ядра та-ламуса нарушения зрительного восприятия были минимальными или отсутствовали. Поражения средней и задней нижневисочной области не привели к нарушениям зрительного поля или снижению зрительной чувствительности (Mishkin, Weiskrantz, 1959; Cowey, Weiskrantz, 1965; Bender, 1973). В то же время, ампутация престриар-ной коры вызвала нарушения остроты зрения и зрительного поля, хотя задачи на зрительное различение в этом случае выполнялись лучше, чем у обезьян с резекцией нижневисочной области.
Исследования зрительного различения на обезьянах отличаются от задачи распознания знакомых незамаскированных объектов, которое нарушается у пациентов со зрительной агнозией объектов конвенционального типа. В задачах на зрительное различение животное должно выучить, что изображение круга на ширме означает, что за ней находится пища, а треугольник означает отсутствие пищи. Это трудная задача для нормальных животных, и для закрепления ее правильного решения требуется от 100 до 200 попыток. После билатеральной ампутации
нижневисочной области у обезьян число попыток, необходимое для выполнения задачи, возрастает до 900-1000. Итак, по-видимому, нижневисочная область участвует в процессе обучения переработке новой зрительной информации, сигнализирующей о наличии пищи. Это обучение направлено преимущественно на сравнение довольно простых сигналов, таких как круг и треугольник, с их моделью памяти, отражающей наличие пищи.
Кроме того, важная роль процесса научения в экспериментальной ситуации заметна в исследованиях реакций одиночных нейронов НВ (нижневисочной) области, особенно ПНВ (передней нижневисочной) области обезьян на объекты и их части (Tanaka et al., 1990; Fujita et al., 1992), а также на ранее не знакомые объекты в разных ракурсах (Logothetis et al., 1995). Например, исследование Логотетиса и др. включало в себя обучение обезьян реагировать на целевой объект или на отвлекающий стимул нажатием на правый или левый рычаг соответственно (Logothetis et al., 1995). В экспериментах Танаки и др. так же, как и во многих похожих экспериментах, для изучения роли процесса научения проводилось многократное предъявление одного и того же стимула (Tanaka et al., 1990).
Между тем, конвенциональный процесс распознания незамаскированных объектов является хорошо усвоенным процессом и не требует обучения в ходе распознания. В основе этого конвенционального процесса могут лежать операции, связанные с функционированием затылочной доли. Ее функционирование нарушается преимущественно у пациентов с конвенциональной зрительной агнозией в результате поражений затылочной доли, которые во многих случаях, особенно при ассоциативной агнозии, также распространяются на задненижнюю область височной доли.
Роль поражений задненижнейи средней областей височнойдоли. Нарушения переработки неконвенциональной информации без признаков конвенциональной зрительной агнозии наблюдаются у пациентов с поражениями задненижней и средней области височной доли, не затрагивающими затылочную долю. Такие случаи редко описываются в литературе, но в одной работе однозначно демонстрируется справедливость этого заключения. Кок в своем исследовании 5 пациентов с опухолями височной доли не наблюдала расстройства распознания объектов в конвенциональных условиях при неограниченном времени предъявления (Кок, 1967). Ограничение времени предъявления объектов на экране тахистоскопа привело к значительному возрастанию порогов распознания объектов. В этих 5 случаях средние и задние участки средней и нижней области височной доли правого или левого полушария были поражены опухолью, а затылочная доля и передняя область височной доли оставались не затронутыми. В случае 4 — мальчик 14 лет с припадками в анамнезе — обнаружены признаки неконвенциональной зрительной агнозии объектов, хотя распознание объектов в конвенциональных условиях было сохранным. Пациент умер после хирургического вмешательства, направленного на лечение припадков. Вскрытие показало наличие обызвествленных масс в нижнесредней области правой височной доли, распространяющихся на нижнезадние области этой доли (рис. 2.1.12).
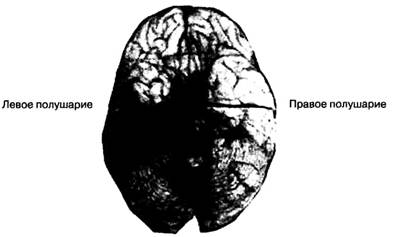
Рис. 2.1.12. Результаты вскрытия в случае 4 - изображение срединной поверхности мозга после удаления окаменелой массы 3,5 х 3,0 х 2,5 см на правой стороне интерпедункулярной цистерны
Масса давила на внутреннюю часть правой нижней области височной доли, затрагивая ее среднюю и частично заднюю часть. Затылочная доля оставалась сохранной. У пациента не наблюдалось признаков конвенциональной зрительной агнозии объектов, но при тестировании с помощью тахистоскопа время распознавания объектов значительно увеличилось (Кок, 1967).
Роль поражений передне-нижней области височной доли.В то же время, поражения передненижней области височной доли не вызывают ни конвенциональной, ни неконвенциональной агнозии объектов, что противоречит результатам исследований реакции одиночных нейронов передневисочной области на конкретные объекты или их части. Показано (Меерсон, 1986), что односторонняя резекция передней области височной доли при лечении эпилепсии лишь иногда вызывает минимальные расстройства выделения и распознания объектов на фоне «зрительного шума» и объектов, предъявленных на экране тахистоскопа при времени экспозиции от 1 до 1000 мс. У пациентов с поражениями затылочной доли среднее время распознания составляло 102 ±15 мс, у здоровых испытуемых контрольной группы — 2,0 ± 0,02 мс, у пациентов после одностороннего хирургического удаления передней области височной доли, включая гиппокамп — 2,2 ± 0,04 мс (рис. 2.1.13 А, Б).
В группе с поражениями передненижней области височной доли (после хирургического лечения эпилепсии височной доли) обнаружена сохранность распознания объектов при уменьшении времени предъявления и серьезные нарушения различения углов, в то время как в группе пациентов с затылочными поражениями наблюдаются серьезные нарушения по обоим тестам. Возможно, это говорит о том, что роль передненижней области височной доли в развитии зрительной агнозии объектов является спорной, несмотря на наблюдающиеся у пациентов серьезные трудности сравнения простых зрительных свойств.
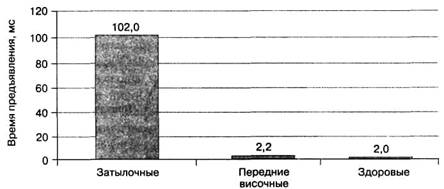
Рис. 2.1.13 А. Пороги времени предъявления, необходимого для распознания объектов пациентами
Дата добавления: 2015-09-18; просмотров: 1378;