ГЛАВА 12. кетовалерил-СоА, который в свою очередь может превращаться в 3-гидроксивалерил-СоА, включаемый в сополимер (рис
кетовалерил-СоА, который в свою очередь может превращаться в 3-гидроксивалерил-СоА, включаемый в сополимер (рис, 12.23),
ЗАКЛЮЧЕНИЕ
Бактерии можно не только использовать как «фабрики» для синтеза белков типа рестриктаз, но и получать с их помощью новые продукты, изменяя метаболизм бактериальных клеток введением в них чужеродных генов или модификацией уже существующих. Можно создавать рекомбинантные микроорганизмы, способные синтезировать самые разные низкомолекулярные соединения: L-аскорбиновую кислоту, краситель индиго, аминокислоты, антибиотики, мономерные единицы различных биополимеров. Общая стратегия при этом состоит во введении в организм хозяина специфических генов, клонированных в подходящем векторе, которые кодируют один или несколько ферментов, катализирующих не свойственные микроорганизму метаболические реакции или влияющих на осуществляемый им в норме биосинтез определенных соединений. По имеющимся данным, создание новых метаболических путей не является технически неосуществимым. Этот подход поможет создать необычные, более эффективные пути синтеза самых разных соединений.
ЛИТЕРАТУРА
Anderson S., С. В. Marks, R. Lazarus, J. Miller, К. Stafford, J. Seymour, D. Light, W.Rastetter, D. Estell.1985. Production of 2-keto-L-gulonate, an intermediate in L-ascorbate synthesis by a genetically modified Erwtnta herbicola. Science230: 144-149.
Bailey L E.1991. Toward a science of metabolic engineering. Science252: 1668-1675.
BerryA. 1996. Improving production of aromatic compounds in Escherichia colt by metabolic engineering. Trends Biotechnol, 14:250-256.
Brooks J. E., F. D. Nathan, D. Landry, L. А.Sznyter, P. Waite-Ress, C. L. Ives, L. S. Moran, B. E. Slatko, J. E. Benner.1991. Characterization of the cloned Bamiil restriction modification sys-
tem: Its nucleotide sequence, properties of the methylase, and expression in heterologous hosts. Nucleic Acids Rex. 19:Я41-850.
Cohen C., D. Shiftman, M. Mcvarcch, V. Aharonowitz.1990. Microbial isopenicillin N synthase genes: structure, function, diversity and evolution. Trends Biotechnol. 8: 105—111.
della-Cioppa G., S. J. Ganger, G. G. Sverlow, T. H. Turpen, L. K. Grill.1990. Melanin production in Escherichia coli from a cloned tyrosinase gene. Bio/Technology 8: 634-638.
Ensley B. D., B. J. Ratzkin, T. D. Osslund, M. J. Simon, L. P. Wackelt, D. T.Gibson. 1983. Expression of naphthalene oxidation genes in Escherichia coli results in the biosynthesis of indigo. Science 222: 167-169.
FloresN., J.Xiao, A. Berry, F. Bolivar, F. Vallc.19%. Pathway engineering for the production of aromatic compounds in Escherichia colt. Nat. Biotechnol. 14:620-623.
Floss H. G. 1987. Hybrid antibiotics—the contribution of the new gene combinations. Trends Biotechnol. 5: 111-115.
Fu J.-F., Y.-H. Tseng. 1990. Construction of lactose-utilizing Xanthomonas campestris and pro-uiiidor. of >.u.ih.4; £.nr;: iVom whey, Лр;>;. Environ. MicrobioL 56:919-923.
Hahn S. K., Υ. Κ. Chang,S. Y. Lee.1995. Recovery and characterization of poly(3-hydroxybutyric acid) synthesized in Alcaligenes eutropfius and recombinant Escherichia coli. Appl, Environ. MicrobioL 61:34-39.
Нillemann D., A. Puhler, W. Wollenen. 1УУ1. Gene disruption and gene replacement in Streptomyces via single stranded DNA transformation of integration vectors. Nucleic Acids Res. 19:727-731.
Hopwood D. A., M. J. Bibb, C. J.Bruton, K. F. Chater, J. S. Fcitelson, J.A. Gil.1983. Cloning Streptomyces genes for antibiotic production. Trends Biotechnol. 1:42-48.
Hopwood D.A., F.Malpartida, H. M. Kicser, H. Ikeda, J.Duncan, I. Fujii, B. A. M. Rudd, H. G. Floss, S. Omura.1985. Production ofhybrid antibiotics by genetic engineering. Nature 314: 642-644.
Howard K. A., C. Card, J. S. Benner, H. L. Callahan, R. Maunus, K. Silber, G. Wilson, J. E. Brooks.1986. Cloning the Dde\ restriction-
Использование рекомбинантных микроорганизмов для получения коммерческих продуктов 273
modification system using a two-step method. Nucleic Adds Res. 14: 7939-7951.
Hutchirtson С R., H. Decker, K.Madduri, S. L. Otten, L. Tang.1993. Genetic control of polyketide biosynthesis in the genus Streptomyces. Anîonie Leeuwcnhfjt'kM: 165—176.
Hutchinson С. R.1994. Drug synthesis by genetically engineering microorganisms. Bio/Technology 12: 375-380.
Hutchinson C. R-, I. FujiL1995. Polyketide synthase gene manipulation: a structure-function approach in engineering novel antibiotics. Anna. Rev. Microbiol. 49: 201-238.
Iked M., K.Nakanishi, K. Kino, R. Katsumata.1994. Fermentative production of tryptophan by a stable recombinant strain of Cotynebacterium glutatnicum with a modified serine-biosynthetic pathway. B'ioscL B'toîechnol. Biochem. 58: 674-678.
Ischida M., K. Miwa, S. Nakamori, K. Sano.December 1989. Process for producing L-trypto-phan. U.S. patent 4, 885, 245.
Isogai T., M. Fukagawa, I. Aramori, M. Iwami, H. Kojo, T. Ono, Y. Ueda, M. Kohsaka, H.Imanaka. 1991. Construction of a 7-aminocephalosporanic acid (7ACA) biosynthetic operon and direct production of 7ACA in Acremonium chtysogenum. Bio/Technology 9: 188-191.
Katz L., S. Donadio.1993. Polyketide synthesis: prospects for hybrid antibiotics. Annu. Rev. Microbiol. 47:875-912.
Kleinkauf H., H. von Dohren.1990. Antibiotics—cloning of biosynthetic pathways. FEBS Lett. 268:405-407.
Krämer R.1996. Genetic and physiological approaches for the production of amino acids. J. Biotechnol. 45: I-2 L
Lazarus R. A., M. Hurle,S. Anderson, D. B. Powers.December 1994. Enzymes for the production of 2-keto-L-gulonic acid. U.S. patent 5. 376, 544.
Lee S. Y., H. N. Chang, Y. K. Chang.1994. Production of poly(ß-hydroxybutyric acid) by recombinant Escherichia со//. Ann. N.Y. Acad. Sei. 721: 43-53.
Lee S. Y., K. S, Yim, H. N. Chang, Y. K. Chang.1994. Construction of plasmids, estimation of plasm id stability, and use of stable plasmids for the production of poly{ß-hydroxybutyric acid)
by recombinant Escherichia со//. J. Biotechnol. 32:203-211.
Lee S. Y., H. N. Chang.1995. Production of poly(3-hydroxybutyric acid) by recombinant Escherichia coti strains: genetic and fermentation studies. Can. J. Microbiol. 41: 207-219.
Magneto S. K., D. L. Lecnutaphong, J. A. DcModcna, J. E.Curtis, J. E. BailOey, J. 1., Galazzo, D. E. Hughes.1991. Actinorhodin production by Streptomyces coelicolor and growth of Streptomyces lividans are improved by the expression of a bacterial hemoglobin. Bio/Technology 9:473-476.
Martin J. F.1987. Cloning of genes involved in penicillin and cephalosporin biosynthesis. Trends Biotechnol. 5: 306-308.
Mi-Daniel R., S. Ebert-Khosla, D. A. Hopwood, C. Khosla.1995. Rational design of aromatic polyketide natural products by recombinant assembly of enzymatic subunits. Nature 375: 549-554.
Mcrmod N., S. Harayama, K. N. Timmis.1986. New route to bacterial production of indigo. Bio/Technology 4: 321-324.
Ozaki A., R. Katsumata, T. Oka.October 1989. Process for producing tryptophan. U.S. patent 4, 874, 698.
Piekaruwicz A., R. Yuan, D. C. Stein.1991. A new method for the rapid identification of genes encoding restriction and modification enzymes. Nucleic Acids Res. 19: 1831-1835.
Rhic H. G., D. Dennis.1995. Role of fadR and otoC(Con) mutations in poly{3-hydroxybutyrate-co-3-hydroxyvalerate) synthesis in recombinant pha + Escherichia coli. Appl. Environ. Microbiol. 61:2487-2492.
Salerno A. J., I. Goldberg. 1993. Cloning, expression, and characterization of a synthetic analog to the bioadhe-sive precursor protein of the sea mussel Mytilus edulis. Appl. Microbiol. Biotechnol. 39:221-226.
Schwarzer Α., A. Puhler.1991. Manipulation of Corynebacteriwn glutamicum by gene disruption and replacement. Bio/Technology 9: 84—87.
Sikora L, A.January 1991. DNA fragment encoding a rubber polymerase and its use, U.S. patent 4. 983, 729.
Stachelhaus T., A. Schneider, M. A. Marahiel.1995. Rational design of peptide antibiotics by targeted replacement of bacterial and fungal domains, Science 269:69-72.
274 ГЛАВА 12
Strausberg R. L., R. P. Link.1990. Protein-based
medical adhesives. Trends ftiotecfinol. 8: 53—57.
Terasawa M., M. Fukushima, Y. Kurusu, H. Yukawa.1990. L-Tryptophan production by the application of highexpressed tryptophanase in Escherichia coli. Process Biochem. In}. 25:172-175.
Walder R. Y., J. L. Hartley, J. E. Dondson, J. A. Wälder.1981. Cloning and expression of the Pstl restriction-modification system in Escherichia coli. Proc. Natl. Acad. Sei. USA 78: 1503-1507.
Weber J. M., J. O. Leung,S. .1.Swanson, К. B. Idler, J. B. McAlfHne. 1991.An erythromycin derivative produced by targeted gene disruption inSaccharopolyspora crythracea. Science252: 114-117.
Yim K. S., S. Y. Lee, H. N. Chang. 1996.Synthesis of poly(3-hydroxybutyrate-co-3-hydroxyvaler-ate) by recombinant Escherichia coli. Biotechnol. Bioeng. 49:495-503.
КОНТРОЛЬНЫЕ ВОПРОСЫ
1. Опишите стратегию выделения гена эндонуклеазы рестрикции EcoRI.
2. Опишите стратегию клонирования гена 2,5-DKG-редуктазы Corynebacterium в Erwinia. Почему это представляет интерес?
3. Предложите стратегию повышения коммерческой ценности клонированного гена 2,5-DKG-редуктазы.
4. Как синтезировать индиго в E. coli?
5. Как повысить количество триптофана, синтезируемого Corynebacterium glutamicum?
6. Предложите стратегию выделения генов, участвующих в биосинтезе антибиотика ундецилпродигиозина, который обычно синтезируется в Streptomyces coelicolor.
7. В чем состоит трудность трансформации различных разновидностей Streptomyces? Как ее преодолеть?
8. Как с помощью генной инженерии увеличить продукцию антибиотика данным штаммом Streptomyces?
9. Опишите один из подходов к созданию модифицированных вариантов неароматических поликетидных антибиотиков типа эритромицина.
10. Как адгезивный белок, обычно синтезируемый мидией Mytitus edulis, можно синтезировать в E. coli?
11. Опишите схему синтеза поли(3-гидроксимасляной кислоты) в E. coli.
12. Что такое сыворотка? Какие важные в промышленном отношении соединения можно из нее получить и каким образом?
ГЛАВА 13.
Биодеградация токсичных соединений и утилизация биомассы
Еще относительно недавно ни у кого не возникало сомнения в том, что окружающая среда — воздух, земля и вода — всегда будут эффективно «перерабатывать» бытовые, промышленные и сельскохозяйственные отходы. Теперь мы знаем, что это не так. Человечество столкнулось с двумя фундаментальными проблемами: переработкой отходов, постоянно образующихся в огромном количестве, и разрушением токсичных соединений, десятилетиями накапливавшихся на свалках, в воде и почве. Правительства разных стран пытались решить эти проблемы законодательным путем, однако к успеху это не привело.
В настоящее время проходит проверку целый ряд технологических, в том числе и биотехнологических, подходов, с помощью которых, возможно, удастся перерабатывать большие количества отходов (например, лигноцеллюлозы) и токсичные вещества. Предпринимаются попытки поощрять те предприятия, которые перерабатывают отходы производства и повторно используют содержащиеся в них полезные вещества.
Употребляемый нами здесь термин «биодеградация» относится к процессу разрушения отходов, попавших в окружающую среду, с помощью живых микроорганизмов, а термин «биомасса» — ко всей совокупности веществ и материалов — побочных продуктов пищевой и перерабатывающей промышленности, — которые раньше считались отходами, а теперь могут служить сырьем для производства многих экономически важных продуктов.
Деградация ксенобиотиков с помощью микроорганизмов
Проблема утилизации токсичных отходов сейчас стоит очень остро. В 1985 г. мировое производство лишь одного из загрязняющих окружающую среду химических веществ, пентахлорфенола, составило более 50 000 т. Раньше токсичные вещества разрушали, сжигая их или обрабатывая другими химикатами, однако это тоже приводило к загрязнению окружающей среды, а кроме того, обходилось очень дорого. В середине 1960-х гг. были обнаружены почвенные микроорганизмы, способные к деградации ксенобиотиков (неприродных, синтетических химических веществ; от греч. xenos, чужой) — гербицидов, пестицидов, хладагентов, растворителей и т. д. Это открытие подтвердило правильность предположения о том, что микроорганизмы можно использовать для экономичного и эффективного разрушения токсичных химических отходов.
Основную группу почвенных микроорганизмов, разрушающих ксенобиотики, составляют бактерии родя Pseudomonas. Биохимические исследования показали, что разные штаммы Pseudomonas способны расщеплять более 100 органических соединений. Нередко один штамм использует в качестве источника углерода несколько родственных соединений.
В биодеградации сложной органической молекулы обычно участвуют несколько разных ферментов. Кодирующие их гены могут иметь хромосомную локализацию, но чаще входят в состав крупных (50—200 т. п. н.) плазмид (табл. 13.1), а
276 ГЛАВА 13
иногда обнаруживаются как в хромосомной, так и в плазмидной ДНК.
Бактерии, разрушающие негалогенированные ароматические соединения, как правило, превращают их в катехол (рис. 13.1) или протокатехоат (рис, 13.2), а затем, в ходе нескольких реакций окислительного расщепления, — в ацетил-СоА и сукцинат (рис. 13.3) или пируват и ацетальдегид (рис. 13.4). Эти последние соединения метаболизируются практически всеми микроорганизмами, Галогенированные ароматические соединения, основные компоненты большинства пестицидов и гербицидов, с помощью тех же ферментов разрушаются до катехола, протокатехоата, гидрохинона или их галогенированных производных, причем скорость их деградации обратно пропорциональна числу атомов галогена в исходном соединении. Дегалогенирование (отщепление замещающего атома галогена от органической молекулы), необходимое для детоксикации соединения, часто осуществляется в ходе неспецифической диоксигеназной реакции, путем замещения галогена в бензольном кольце на гидроксильную группу. Эта реакция может происходить как в ходе биодеградации исходного галогенированного соединения, так и потом.
Метаболические пути биодеградации ксенобиотиков, созданные методами генной инженерии
Некоторые микроорганизмы обладают природной способностью к деградации различных ксенобиотиков, однако следует иметь в виду, что: I ) ни один из них не может разрушать все органические соединения; 2) некоторые органические соединения в высокой концентрации подавляют функционирование или рост деградирующих их микроорганизмов; 3) большинство очагов загрязнения содержит смесь химикатов, и микроорганизм, способный разрушать один или несколько ее компонентов, может инактивироваться другими компонентами; 4) многие неполярные соединения адсорбируются частицами почвы и становятся менее доступными; 5) биодеградация органических соединений часто происходит довольно медленно. Часть этих проблем можно решить, осуществив конъюгационный перенос плазмид, которые кодируют ферменты разных катаболических путей, в один реципиентный штамм (рис. 13.5). Если две плазмиды содержат гомологичные участки, то между ними может произойти рекомбинация с образованием гибридной плазмиды, которая имеет больший размер и обладает свойствами исходных плазмид. Если же две плазмиды не содержат гомологичных участков и относятся к разным группам несовместимости, то они могут сосуществовать в одной бактерии.
Перенос плазмид
В 1970-х гг. Чакрабарти и его коллегами был создан первый бактериальный штамм, обладающий более широкими катаболическими возможностями. Он расщеплял большинство углеводородов нефти и был назван «супербациллой». Для его получения использовали плазмиды, каждая из которых кодировала фермент, расщепляющий определенный класс углеводородов: плазмида САМ детерминировала деградацию камфары, ОСТ — октана, NAH — нафталина, XYL — ксилола (рис. 13.5). Сначала путем конъюгации пере-
| Таблица 13. 1. Плазмиды Pseudomonas, их размер и соединения, за разрушение которых ответственны кодируемые ими ферменты1) | |||
| Плазмида2) | Деградируемое соединение | Размер плазмиды, т. п. н. | |
| SAL | Салицилат | ||
| SAL | Салицилат | ||
| SAL | Салицилат | ||
| TOL | Ксилол и толуол | ||
| pJP1 | 2 ,4 - дихлорфеноксиуксусная | ||
| кислота | |||
| pJP2 | 2,4-дихлорфеноксиуксусная | ||
| кислота | |||
| pJP3 | 2 ,4- дихлорфеноксиуксусная кислота | ||
| CAM | Камфара | ||
| XYL | Ксилол | ||
| pAC31 | 3,5-дихлорбензоат | ||
| pAC25 | 3-хлорбензоат | ||
| pWWO | Ксилол и толуол | ||
| NAH | Нафталин | ||
| XYL-K | Ксилол и толуол | ||
| 1) Из работы Cork, Krueger, Adv. Appl. Microbiol. 36: 1-66, 1991, с изменениями. | |||
| 2) Плазмиды с одинаковым названием кодируют ферменты одного и того же катаболического пути, хотя могут быть получены в разных лабораториях и иметь разные размеры. |
Биодеградация и утилизация биомассы 277
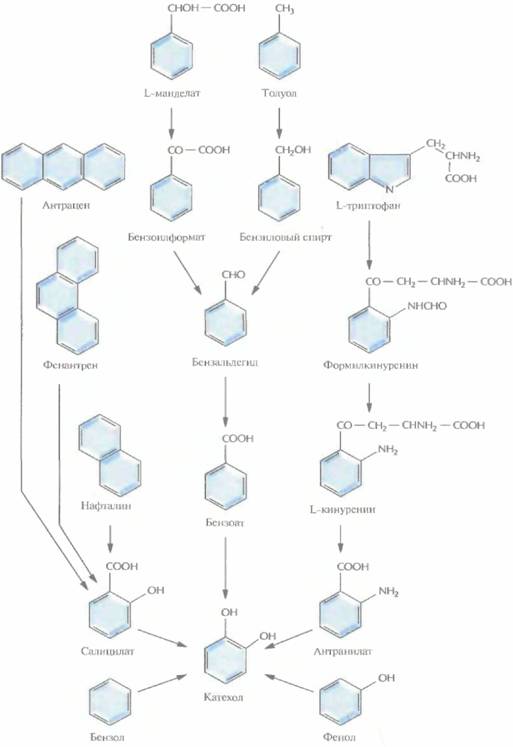
|
| Рис. 13.1. Пути ферментативного превращения ароматических соединений в катехол бактериями, разрушающими ксенобиотики. |
Дата добавления: 2015-07-14; просмотров: 733;