Мембраны цитоплазмы
Общей чертой всех мембран клетки, внешней плазматической мембраны и всех внутриклеточных мембран и мембранных органоидов будет то, что они представляют собой тонкие (6-10 нм) пласты липопротеидной природы (липиды в комплексе с белками), замкнутые сами на себя.
В клетке нет открытых мембран со свободными концами. Мембраны клетки всегда ограничивают полости или участки, закрывая их со всех сторон и тем самым отделяя содержимое таких полостей от окружающей их среды. Так, плазматическая мембрана, покрывая всю поверхность клетки, имеющей сложную форму и многочисленные выросты, нигде не прерывается, замкнута. Она отделяет содержимое цитоплазмы от окружающей клетку среды. Внутриклеточные замкнутые мембраны образуют пузырьки – вакуоли шаровидной или уплощенной формы. В последнем случае образуются плоские мембранные мешки, или цистерны.
Часто полости, отграниченные мембранами, имеют сложную форму, напоминающую губку или сеть; и в этом случае такие полости без перерывов отграничены мембраной. В этих случаях также мембраны разделяют две структурные фазы цитоплазмы: гиалоплазму от содержимого вакуолей и цистерн. Такое же свойство имеют мембраны митохондрий и пластид: они разделяют внутреннее содержимое от межмембранных полостей и гиалоплазмы.
Ядерная оболочка тоже может быть представлена в виде перфорированного полого двойного мембранного мешка шаровидной формы мембраны ядерной оболочки разграничивают, отделяют друг от друга кариоплазму и хромосомы от полости перинуклеарного пространства и гиалоплазмы.
Эти общие морфологические свойства клеточных мембран определяются их химическим составом, их липопротеидной природной.
Все клеточные мембраны имеют ряд общих для них свойств:
1. Их структурную основу составляет двойной слой липидов.
2. В плоскости липидных слоев расположены белковые молекулы.
3. Белки и липиды ассиметрично расположены в плоскости мембран.
4. Белки и липиды обладают латеральной подвижностью в плоскости мембран.
5. Мембраны изменчивы в зависимости от функционального состояния.
6. Мембраны ассоциированы с цитоплазматическими белками, микрофиламентами и микротрубочками посредством специальных белков.
7. Рост мембран происходит путем расширения их поверхности за счет включения нового материала в виде готовых замкнутых пузырьков (везикул).
8. Синтез компонентов и сборка цитоплазматических мембран происходят за счет активности гранулярного эндоплазматического ретикулума.
Количество липидов и белков в большинстве мембран почти одинаковое (40-60%) по массе, но в численном отношении мелких липидных молекул намного больше, чем тяжелых белковых, и разнообразие липидов невелико. Белки же мембран отличаются большим разнообразием. Углеводный компонент (2-10% от веса), является компонентом главным образом плазматической мембраны. К липидам относится большая группа органических веществ, обладающих плохой растворимостью в воде (гидрофобность) и хорошей растворимостью в органических растворителях и жирах (липофильность). Состав липидов, входящих в мембраны клетки, очень разнообразен. Характерными представителями липидов, встречающихся в клеточных мембранах, являются фосфолипиды (глицерофосфатиды), сфингомиелины и из стероидных липидов – холестерин.
Характерной особенностью липидов мембран является разделение их молекулы на две функционально различные части: неполярные, не несущие зарядов хвосты, состоящие из жирных кислот, и заряженные полярные головки, которые несут на себе отрицательные заряды или могут быть нейтральными. Если полярные липиды смешать с водой, то образуется эмульсия, состоящая из мицелл. При этом незаряженные (гидрофобные) хвосты будут стремиться образовывать однородную фазу в центре мицеллы, и заряженные, гидрофильные, головки будут торчать в водную фазу.
Смешивая с водой экстрагированные из мембран липиды или беря смеси разных липидов, можно получить бимолекулярные слои или мембраны, где периферические зоны слоя, смотрящие в водную фазу, будут содержать исключительно полярные головки, а незаряженные хвосты будут образовывать общую гидрофобную центральную зону такой образовавшейся мембраны. Эта способность липидов самопроизвольно образовывать мембранные структуры определяется свойствами самих липидов.
Молекулы липидов в искусственных слоях или в составе клеточных мембран очень подвижны, они могут перемещаться в плоскости мембран с высокой скоростью. Но перескок из одного слоя в другой – редкое состояние. С таким текучим слоем связаны мембранные белки.
В среднем в липопротеидных мембранах белки по весу составляют 50%. Но количество белков в разных мембранах может быть различным. Так в мембранах митохондрий на долю белков приходится около 75%, а в плазматической мембране клеток миелиновой оболочки - около 25%. Но так как липидные молекулы имеют небольшой размер (около 0,5 нм) и молекулярный вес, их число по отношению к числу белковых молекул выше в 50 раз. Поэтому белковые молекулы как бы вкраплены в билипидный слой мембраны.
Часть из них связана с липидными головками с помощью ионных (солевых) связей и поэтому легко экстрагируется из мембран растворами солей. Другие образуют солевые связи с полярными участками липидов через взаимодействие с ионами Mg2+ или Ca2+, такие белки экстрагируются с помощью хелатных соединений. Такие легко экстрагируемые белки большей частью расположены на мембранах со стороны цитоплазмы. В цитоплазматической мембране эти белки тесно связаны с белковыми структурами цитоскелета.
Большая часть белков взаимодействует с липидами в составе мембран на основе гидрофобных связей. Оказалось, что многие мембранные белки состоят как бы из двух частей: из участков, богатых полярными (несущими заряд) аминокислотами, и участков, обогащенных неполярными аминокислотами (глицином, аланином, валином, лейцином).
Такие белки в липидных слоях мембран располагаются так, что их неполярные участки как бы погружены в “жирную” часть мембраны, где находятся гидрофобные участки липидов. Полярная (гидрофильная) же часть таких белков взаимодействует с головками липидов и обращена в сторону водной фазы, поэтому такие белки, связанные с липидами путем гидрофобных взаимодействий, практически не экстрагируются в водных фазах. Их можно выделить, лишь разрушая мембрану, экстрагируя из нее липиды или органическими растворителями, или детергентами. Поэтому эти белки мембран и называют интегральными.
Размер интегральных мембранных белков в среднем равен 8 нм, но встречаются крупные белки – до 35 нм величиной (белок тилакоидов хлоропластов). Обычно это очень асимметричные по своей природе белки и соответственно асимметрично локализованы в мембране: их разные функциональные части локализованы по обе стороны мембраны, и все белки данного типа расположены одинаково.
С цитоплазматической стороны мембраны интегральные белки связаны с периферическими белками. Часть белков может быть связана только с полярными группами липидов и может находиться на поверхности билипидного слоя; другие белки могут частично или даже полностью погружены из-за гидрофобных свойств своих участков в липидный слой; третьи – могут пронизывать мембрану насквозь. Интересно, что большая часть липидных молекул (70%) не связана с белками, так что белковые молекулы как бы плавают в “липидном озере”.
Липидные молекулы двигаются вдоль липидного слоя, могут вращаться вокруг своей оси, а также переходить из слоя в слой, что происходит редко и с помощью специальных переносчиков.
Белки, плавающие в “липидном озере”, также обладают латеральной, продольной подвижностью, но скорость их перемещения в десятки и сотни раз ниже. Белки плазматической мембраны, гликопротеины, часто имеют олигосахаридные цепочки, смотрящие на внеклеточную среду.
Особенно отличаются мембраны по составу белков, которые, главным образом, определяют функциональные свойства мембран. По биологической роли мембранные белки можно разделить на три группы: ферменты, рецепторные белки и структурные белки.
2. Зарисуйте и подпишите различные модели клеточных мембран:
а) Плоскостную и трехмерную модели мембраны.
| Рис.2. Плоскостное изображение жидкостно-мозаичной модели мембраны. | 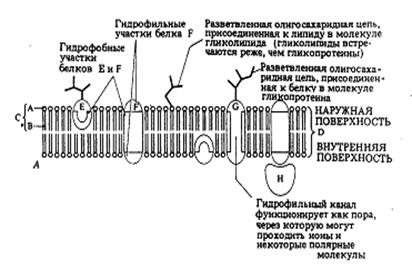
|
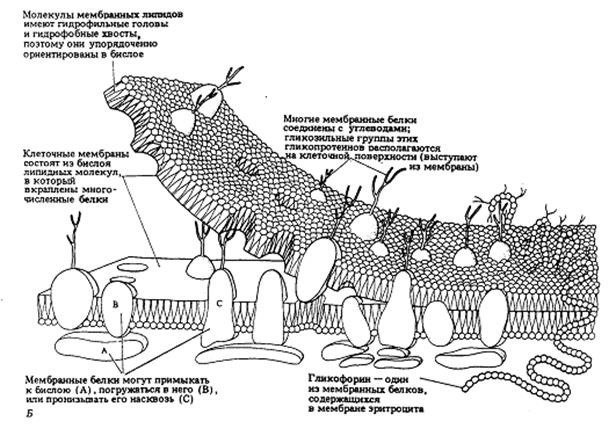 Рис.3. Трехмерная модель мембраны.
Рис.3. Трехмерная модель мембраны.
|
б) Мозаичную модель мембраны.
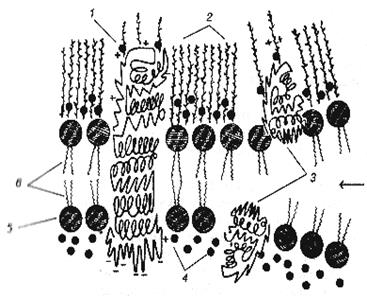
| Рис.4. Мозаичная модель клеточной мембраны и поверхности мембраны: 1 – интегральный белок; 2 – полисахариды гликокаликса; 3 – полуинтегральные белки; 4 – периферический белок; 5 – слой гидрофильных головок липидных молекул; 6 – гидрофобные концы бислоя липидных молекул. |
3. Изучите структуру клеточных мембран.
а) Рассмотрите (рис.5) и зарисуйте взаимодействие белков с липидными слоями.
| Рис.5. Взаимодействие белков с липидными слоями. А - белковая молекула, связанная ионными взаимодействиями; Б, В – гидрофильно (б1, в1) - гидрофобные (б2, в2) взаимодействия белков с липидами (по Покровскому и Тутельяну, 1976). |
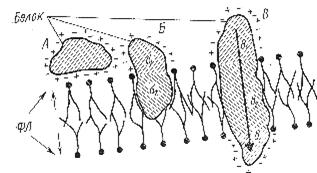
|
б) Рассмотрите (рис.6) и зарисуйте схему строения молекул фосфолипидов.
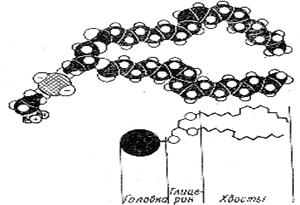
Рис. 6. Схема строения молекул фосфолипидов.
Дата добавления: 2015-08-08; просмотров: 4431;