БИОФИЗИЧЕСКИЕ МЕХАНИЗМЫ ТРАНСПОРТА ВЕЩЕСТВ (МАССОПЕРЕНОСА) ЧЕРЕЗ БИОЛОГИЧЕСКИЕ МЕМБРАНЫ
Лекция № 13
Принято различать пассивный и активный транспорт веществ через клеточные мембраны. К первому из них относят трансмембранный массоперенос, происходящий в направлении действия концентрационного, электрического, осмотического и фильтрационного (гидростатического) градиентов. Так как по отношению к одному и тому же веществу они могут быть направлены противоположно, то при анализе такого массопереноса необходимо учитывать термодинамическое сопряжение всех физико-химических градиентов.
Активным транспортом называют перенос вещества в направлении, противоположном тому, которое предопределено термодинамическим сопряжением перечисленных выше градиентов. Движущей силой активного транспорта служит химический потенциал, обусловливающий в БМ течение ферментативных реакций, поставляющих свободную энергию для преодоления градиентов.
Все вышеперечисленные градиенты имеют неодинаковое значение в переносе веществ через клеточные мембраны.
Массоперенос, происходящий в результате суммарного действия осмотического и гидростатического градиентов, принято называть конвекционным (конвективным) потоком.Им определяется прежде всего трансмембранный ток воды вместе с растворенными в ней веществами.
Кинетика биофизических процессов массопереноса.При исследовании кинетики биофизических процессов измеряют скорости их протекания в различных условиях. При описании любого явления переноса обобщенными координатами скорость процесса выражают в виде производной соответствующей обобщенной координаты (х) по времени:
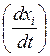
(38)
где х – обобщенная координата − величина, характеризующая перемещение или перенос.Во многих явлениях природы скорости процессов прямо пропорциональны соответствующим обобщенным силам (Xi), характеризующим причины возникновения соответствующего процесса:

(39)
где Аi− коэффициент пропорциональности; Хi − обобщенная сила − величина, характеризующая причину перемещения или переноса.
Кинетика сопряженных процессов массопереноса.Обычно процессы массопереноса происходят под действием сразу нескольких обобщенных сил. Например, при переносе заряженных частиц через клеточную мембрану имеют место и диффузия, и электрический ток. В этом случае происходит так называемое термодинамическое сопряжение между диффузией и электрическим током, и скорость каждого из этих процессов зависит не только от «своей» обобщенной силы, но и от всех обобщенных сил, действующих в системе.
Градиенты концентрации и температуры выполняют роль обобщенных сил, а обобщенными координатами служат масса вещества и количество тепла, переносимые в системе из одной точки пространства в другую. Транспорт вещества в такой системе определяется не только градиентом концентрации, но и градиентом температуры.
Если в какой-то части системы как концентрация вещества, так и температура выше, чем в другой, то grad С и grad Т имеют одинаковые знаки, и процессы, происходящие под действием их, усиливают друг друга и следовательно массоперенос больше, чем в условиях существования только одного из градиентов.
Если же grad С и grad T разнонаправлены (имеют разные знаки) и
grad C<< grad T, то 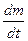 <0, что означает перенос вещества вопреки действию градиента концентрации.
<0, что означает перенос вещества вопреки действию градиента концентрации.
В таком случае под действием разности температур в двух частях системы вещество будет перемещаться из области, где его концентрация низка, в область высокой концентрации. Данное явление называют термодиффузией.Оно, в частности, используется в атомной промышленности − посредством термодиффузии разделяют изотопы, например 235U и 238U. Это возможно потому, что коэффициенты А для этих изотопов урана различны, причем система уравнений позволяет рассчитать те условия, при которых потоки легкого и тяжелого изотопов направлены в противоположные стороны. Для осуществления термодиффузии необходимы значительные затраты свободной энергии. В транспорте веществ через клеточные мембраны термодиффузия отсутствует, поскольку grad Т на мембране равен нулю.
Другим примером может быть система, в которой существуют как градиент концентрации, так и градиент осмотического давления. Такой системой является живая клетка в солевом растворе. Можно подобрать условия, при которых вода пойдет против осмотического градиента, благодаря термодинамическому сопряжению. Так, иногда клетки в культурах тканей при помещении в раствор соли не отдают воду, как требует теория осмоса, а набухают за счет поступления в них воды из окружающего раствора, осмотическое давление которого выше, чем в цитоплазме.
На сильно засоленных почвах живут своеобразные растения − солянки, в корни которых вода поступает вопреки действию градиента осмотического давления. Такие факты ставили биологов в тупик; некоторые ученые-виталисты даже рассматривали их как доказательство существования особой «жизненной силы», не подчиняющейся физическим законам. Использование теории термодинамического сопряжения позволило дать всем этим явлениям как качественное, так и количественное объяснение.
Транспорт веществ через клеточные мембраны также осуществляется благодаря термодинамическому сопряжению разнообразных процессов. Только в тех случаях, когда молекулы транспортируемого вещества не имеют заряда, химически и осмотически инертны, процесс массопередачи сводится к простой диффузии и подчиняется уравнению Фика.

(40)
где АD =D·s , D – коэффициент диффузии, s – площадь слоя мембраны.
Практически такими свойствами обладают в организме только азот и инертные газы; очень приблизительно − кислород и углекислый газ. Во всех остальных случаях (при транспорте воды, солей, углеводов, жирных кислот, пептидов и других веществ) вместе с концентрационным градиентом действуют и другие.
Например, при транспорте многих ионов идут, по меньшей мере, два процесса: диффузия вещества и перенос заряда, а значит, взаимодействуют концентрационный и электрический градиенты.
Сопряженный массоперенос заряженных частиц (ионов) через биологическую мембрану.Как уже говорилось, при транспорте ионов через плазматическую мембрану имеет место термодинамическое сопряжение концентрационного и электрического градиентов: grad С и grad U:
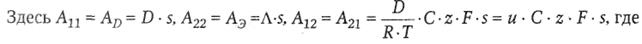
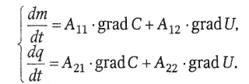 |
D-коэффициент диффузии (м2 · с-1), 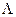 − удельная электропроводность (См · м-1),
− удельная электропроводность (См · м-1),
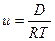
− подвижность (м2 · моль · Дж-1 · с-1),
С − молярная концентрация переносимого вещества (моль · м-3], F − число Фарадея − заряд 1 моля одновалентного иона − F= 9,65 104 Кл · моль-1, z − валентность, s − площадь поверхности массопереноса (м2).
Решая совместно эту систему уравнений, приходим к следующему выражению: 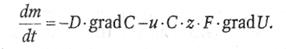 (41)
(41)
В этом уравнении − поток вещества; знаки «минус» в правой части отображают то обстоятельство, что масса вещества при его транспорте убывает там, где она больше.
Перейдем от потока вещества к плотности потока вещества (моль · м-2 · с-1):
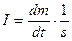
(42)
Эту величину называют также интенсивностью массопереноса:
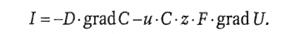 (43)
(43)
Электродиффузионное уравнение Нернста−Планка позволяет рассчитать интенсивность массопереноса в однородных (гомогенных) средах (например, в растворах). Присутствие биологических мембран между растворами (в частности, между цитозолем и интерстицием) искажает результаты таких расчетов, поскольку по физико-химическим свойствам мембраны отличаются от водных растворов солей. Для учета поправок на эту неоднородность введено понятие проницаемости биомембраны.
Проницаемость клеточных мембран.Проницаемостью биологической мембраны называют ее способность пропускать сквозь себя определенные вещества в той или иной степени.Расчет мембранной проницаемости, точнее коэффициента проницаемости (р), проводят по формуле:
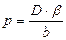
(44)
где b − толщина биомембраны, D − коэффициент диффузии, 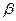 − коэффициент распределения того или иного вещества между водой и липидом:
− коэффициент распределения того или иного вещества между водой и липидом:
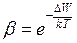
(45)
где 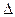 W − разность энергий, которыми обладает частица (молекула) данного вещества в воде и липиде.
W − разность энергий, которыми обладает частица (молекула) данного вещества в воде и липиде.
У ионов эта энергия находится в сильной зависимости от диэлектрической проницаемости (ɛ) сред, в которых они присутствуют, − среда с большей ɛ сильнее взаимодействует с ионами: W=f(ɛ).
В постоянном электрическом поле ɛ воды = 81, а ɛлипида = 2−3. Следовательно, разность энергий ( W), которыми обладает молекула данного вещества в воде и липиде, тем больше, чем больше разница в диэлектрической проницаемости ( ɛ) воды и липида. А поскольку W в формуле входит в показатель степени при основании натуральных логарифмов, то ɛ на влияет очень сильно.
 |
Потенциальный барьер, который нужно преодолеть для переноса 1 иона из интерстиция в бимолекулярный липидный каркас биомембраны, рассчитывают по формуле Борна:
(46)
где z − валентность иона, е − заряд электрона, r − радиус иона, k = 1,38 · 10-23 Дж · К-1 (постоянная Больцмана), T − абсолютная температура.
Расчет свидетельствует, что для переноса через биомембрану одновалентного иона, имеющего радиус 0,2 нм, необходимо затратить энергию ( W), равную 70 · kТ, чему соответствует 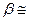 10 -20. Это означает, что переход ионов и других гидрофильных веществ через липидный мембранный каркас невозможен. У гидрофобных веществ на много порядков больше, и они растворяются в мембранных липидах.
10 -20. Это означает, что переход ионов и других гидрофильных веществ через липидный мембранный каркас невозможен. У гидрофобных веществ на много порядков больше, и они растворяются в мембранных липидах.
Поэтому механизмы переноса гидрофильных и гидрофобных веществ через биологические мембраны различаются коренным образом.
Транспорт липофильных веществ через биологические мембраны. В1895 г. Э. Овертон установил, что вещества тем легче проникают в клетку, чем выше их растворимость в липидах, а она высока у неполярных агентов. Липофильные соединения проходят через БМ, растворяясь в ее липидах и двигаясь по законам диффузии в вязкой среде. Такой транспорт зависит от величины и формы проникающих молекул, а также от вязкости мембраны. Естественно, что скорость трансмембранного переноса жирорастворимых веществ испытывает сильную зависимость от температуры. Определив зависимость мембранной проницаемости для тех или иных агентов от температуры (рис. 50), находят энергию активации (Wa) транспорта их через БМ. Оказалось, что величины Wa для этиленгликоля (60 кДж · моль-1), глицерина (77 кДж · моль-1), эритрита (87 кДж · моль-1) близко совпадают с энергией дегидратации этих спиртов. По-видимому, прежде чем войти в липидный слой мембраны, молекулы неэлектролитов подвергаются дегидратации. Только после освобождения от водной оболочки они внедряются в жидкокристаллическую структуру БМ и диффундируют в ней.
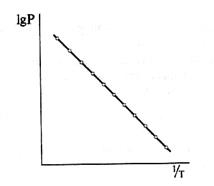
Рис. 50. Зависимость проницаемости биомембраны от обратной температуры: по оси абсцисс — величина, обратная абсолютной температуре; по оси ординат — логарифм проницаемости.
Интересно, что транспорт глицерина через плазмолемму эритроцитов человека и крысы отличается высокой скоростью и гораздо более низкой Wa по сравнению с энергией дегидратации. По-видимому, здесь нет нужды в предварительной дегидратации и, следовательно, глицерин не диффундирует сквозь липидный бислой эритроцитарных мембран человека и крысы. Вероятно, в них присутствуют специи- альные переносчики глицерина.
Молекулярные компоненты клеточной мембраны непрерывно перемещаются в пределах своего слоя (латеральная диффузия), задерживаясь на одном месте около 10-7 с. Поэтому существуют мгновения, когда освободившееся место оказывается еще не занятым соседними молекулами мембраны. Этот процесс аналогичен возникновению «вакансий» в кристаллической решетке. Однако в жидких кристаллах вероятность образования вакансий на несколько порядков выше, чем в твердых.
Молекулы неэлектролитов могут занять под действием физико-химических градиентов возникшую на миг вакансию в липидном слое БМ. Разумеется, такое вторжение в ее жидкокристаллическую структуру возможно только в том случае, если диффундирующая молекула по геометрическим параметрам укладывается в размеры вакантного места (ячейки). Если же калибр проникающих молекул приходится на диапазон от одного до двух диаметров ячейки, то для продвижения на 1 шаг такая молекула должна выждать момент, когда одновременно станут свободными две соседние ячейки, смежные с тем местом, где она находится. Подобное событие совершается значительно реже, чем образование одиночной вакансии, но в жидком кристалле оно вероятно. Еще меньшую, но реальную вероятность имеет одномоментное возникновение трех смежных вакансий по соседству с местонахождением диффундирующей молекулы, размеры которой втрое больше диаметра одиночной ячейки.
Следовательно, чем крупнее частицы проникающего через мембрану вещества, тем меньше вероятность его проникновения сквозь нее (при одинаковых прочих условиях). Это отражается на скорости трансмембранного переноса веществ: крупные молекулы дольше ждут того момента, когда смогут продвинуться на один шаг, и поэтому проходят БМ медленнее. У низкомолекулярных неэлектролитов скорость мембранного транспорта обратно пропорциональна квадратному корню, а у высокомолекулярных − кубическому корню из молекулярной массы. Показателем скорости перемещения веществ через БМ служит коэффициент диффузии (D). Поэтому указанные зависимости проницаемости от молекулярной массы ( 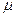 ) обычно выражают так: для мелких молекул (от водорода до трисахаридов) − D 1/2= const, а для более крупных молекул − D 1/3 = const.
) обычно выражают так: для мелких молекул (от водорода до трисахаридов) − D 1/2= const, а для более крупных молекул − D 1/3 = const.
Частицы диффундирующего вещества перемещаются в БМ не непрерывно, а дискретно − скачками (от одного вакантного места к другому, выжидая его образование), в силу чего изложенная схема транспортного процесса получила название гипотезы скачков. Она основана на положениях статистической физики. В зарубежной литературе эта схема фигурирует под названием гипотезы кинков.
Кинками называют лабильные (временно существующие) и непрерывно перемещающиеся структурные дефекты в углеводородной части БМ. Ими определяется наличие в мембране свободных объемов (вакансий, временных дырок). Согласно гипотезе кинков, в такую вакансию могут встраиваться мелкие молекулы неэлектролитов (в том числе ассоциаты воды) и мигрировать вместе с ней. Поэтому о кинках говорят как о «кинетических каналах» мембраны. Коэффициент диффузии кинков вместе с захваченными ими молекулами проникающего вещества довольно высок − до 10-9 м2 · с-1. Следовательно, кинки обеспечивают быструю диффузию.
Транспорт гидрофильных веществ через биологические мембраны.Гидрофильные вещества не способны перейти из водного раствора (интерстиция или цитозоля) в липидный каркас клеточной мембраны. Для трансмембранного переноса у них есть две возможности:
1) . одеться в гидрофобный «чехол» и в таком виде раствориться в липидной фазе мембраны (наподобие транспорта ионов калия валиномицином) − так происходит транспорт гидрофильных веществ при помощи переносчиков;
2) . пройти через такие места в биомембране, где ε велика; причем, такую же величину, как в интерстиции и в цитозоле, ε имеет в сквозных порах, заполненных водой, т. е. в мембранных каналах − так происходит транспорт гидрофильных веществ по каналам в биомембране.
Транспорт с участием переносчиков.Транспорт многих гидрофильных веществ (моносахаридов, аминокислот, некоторых ионов и др.) обеспечивают подвижные переносчики. В 1971 г. группа американских ученых обнаружила в БМ пептид, имеющий циклическую структуру и способный селективно переносить ионы. Преимущества транспорта гидрофильных веществ при помощи переносчиков по сравнению со свободной диффузией через липидный бислой клеточной мембраны иллюстрирует перенос глюкозы из плазмы крови в эритроцит. У глюкозы пять гидроксильных групп, способных образовывать водородные связи. Теоретически Wa переноса глюкозы из водного раствора в гидрофобную область БМ должна составлять около 80 кДж · моль-1. В эксперименте Wа транспорта глюкозы через эритроцитарную мембрану значительно меньше. Облегчение трансмембранного переноса обусловлено тем, что глюкоза преодолевает плазмолемму не путем свободной диффузии через липидный бислой, а при помощи переносчика. Поэтому движение глюкозы сквозь мембрану эритроцита происходит примерно в 10 тыс. раз быстрее, чем можно было ожидать при ее свободной диффузии в мембранных липидах.
Из эритроцитов человека выделен специфический переносчик глюкозы, оказавшийся интегральным белком плазмолеммы. Искусственные липидные мембраны, в которые его встраивают, приобретают селективную проницаемость, свойственную клеточным мембранам человека и животных: через них с большой скоростью переносится только D-глюкоза, тогда как L-глюкоза практически не проникает.
В плазматической мембране бактериальной клетки − кишечной палочки (Е. coli) обнаружено несколько десятков транспортных белков, каждый из которых переносит определенное вещество, причем для каждого из сахаров есть специфический переносчик.
Переносчики в БМ могут работать, используя разные способы перемещения (миграционный, ротационный, сдвиговый и т. д.). Среди мигрирующих переносчиков можно выделить две разновидности. Одни транспортеры мигрируют внутри мембраны, взаимодействуя с переносимым веществом только на ее поверхностях. Этот механизм транспорта называют малой каруселью. Другие мигрирующие переносчики способны покидать биомембрану и выходить в примембранное пространство в поисках транспортируемого агента. Поиск направляется действием электростатических сил и химическим взаимодействием. Вместе с переносимым веществом транспортер второго типа возвращается в БМ, проходит ее насквозь, выходит в противоположное примембранное пространство и оставляет там свой «багаж». В этом случае говорят о большой «карусели». Тип «карусели» зависит от поверхностно-активных свойств и растворимости самого переносчика. По механизму малой карусели работают транспортеры, плохо растворяющиеся в воде и являющиеся поверхностно-активными веществами.
Миграционный механизм присущ переносчикам, размеры которых меньше, чем толщина БМ (рис. 51, а). Вместе с тем транспортерами могут служить крупные белковые молекулы и их комплексы, прободающие насквозь липидный бислой. Они переносят вещества через БМ посредством ротации или сдвига на расстояние,
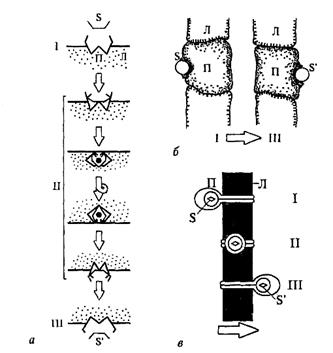 |
равное толщине мембраны.
Рис. 51. Схема работы мембранных переносчиков разных типов: а − миграция; б − ротация; в − сдвиг. S − переносимое вещество до переноса, S' − переносимое вещество после переноса; П − переносчик; Л − липидные компоненты мембраны; I − состояние до начала переноса; II − процесс переноса; III − состояние после переноса.
Ротационный механизм (рис. 51, б) заключается в перевороте крупной молекулы переносчика вокруг оси, лежащей в плоскости, мембраны, в результате чего транспортируемое вещество, посаженное на один конец такой молекулы, оказывается на противоположной стороне БМ. Ротационный перенос требует значительных затрат энергии и может оказаться эффективным только в том случае, если 1 молекула переносчика транспортирует одномоментно (за один поворот) много молекул переносимого вещества.
Более выгодным в энергетическом отношении является механохимический процесс в молекуле переносчика, заключающийся не в полном ее перевороте, а в сдвиге отдельных областей относительно неподвижной части, причем вместе с участком переносчика, уходящим с поверхности в глубь мембраны, в нее погружается и транспортируемое вещество (рис. 51, в). Это напоминает движение ленты транспортера.
По-видимому, в разных БМ и при транспорте различных веществ работают разнообразные переносчики: одни мигрируют сквозь мембрану, другие − переворачиваются, третьи − претерпевают конформационные перестройки со сдвигом активных, т. е. взаимодействующих с транспортируемым веществом групп относительно неподвижных.
Перспективным направлением в изучении этой проблемы является выяснение «машинных» свойств белковой молекулы (Блюменфельд, 1977). Говоря о молекулярных машинах живой клетки, Л.А. Блюменфельд стремился учесть не только статистические, но и механические принципы работы биологических молекул. Характерной особенностью белков является существование кинетической неравновесности на уровне вторичной и третичной структур, в силу чего многие химические реакции, в которые вступают белковые молекулы, сопровождаются конформационными перестройками (изменениями геометрической формы молекулы за счет свободного вращения отдельных ее фрагментов относительно простых углерод-углеродных связей). Такие изменения вторичной и третичной структур белковой молекулы приводят к изменению ее пространственных координат, т. е. к механическому перемещению. Молекула переносчика, вступив в реакцию с транспортируемым веществом и посадив его на себя, испытывает конформационные превращения, выражением чего может быть тот или иной механизм перемещения ее в биомембране.
Транспорт по мембранным каналам.Гипотеза о существовании в живых тканях пор, заполненных водой, по которым осуществляется массоперенос, была сформулирована Э.В. Брюкке в 1842 г. при исследовании транспорта воды через стенку мочевого пузыря. Однако только через столетие − во второй половине XX в. − гипотеза Брюкке стала общепринятой концепцией, чему способствовали работы Ходжкина и Хаксли (1952), Девсона и Даниелли (1955) и исследования свойств грамицидинового канала, встроенного в искусственный липидный бислой (Хладни, Хайдон, 1970; 1972).
Согласно современной концепции, мембранный канал представляет собой интегральный белок (белковый комплекс, или гликопротеид), встроенный в липидный бимолекулярный каркас мембраны, пронизывающий ее насквозь и обеспечивающий перенос веществ через нее в сторону более низкого электрохимического потенциала. Вторичная структура компонентов мембранного белкового комплекса имеет характер β-складчатости с цилиндрической порой внутри, заполненной водой. Коэффициент проницаемости ионных каналов составляет 10-8−10-9 м · с-1, что на 5−6 порядков меньше скорости переноса ионов при свободной диффузии в водной среде. Замедление обусловлено тем, что движение иона по каналу представляет собой последовательное замещение молекул воды гидратной оболочки иона на полярные группы, выстилающие полость канала.
Хилле в 1977−1984 гг. предложил функциональную модель ионного канала, согласно которой в нем имеются два основных компонента: селективный фильтр и воротный механизм. Первый имеет жесткую структуру, т. е. в этой части белковый комплекс, образующий канал, не может изменять размеры поры внутри него и регулировать проницаемость мембраны. Функция селективного фильтра − пропускать через канал определенное вещество или группу сходных с ним веществ, т. е. отбирать их из других.
Регулирование мембранной проницаемости обеспечивается так называемыми воротными процессами. Они осуществляются «воротами канала», которые представляют собой части белкового комплекса, способные «раскручиваться» и «скручиваться» в ходе их механохимических реакций и благодаря этому создавать просвет внутри белкового комплекса или перекрывать его (сжимать или восстанавливать пору).
Профиль просвета имеет размер, соизмеримый с размером пропускаемого вещества (десятые доли нанометра). Поэтому движение веществ по каналу однорядное и, следовательно, канал может пребывать только в одном из двух противоположных состояний: закрытом или полностью открытом, при котором достигается максимальная скорость транспорта. Например, скорость движения натрия и кальция через соответствующие потенциалзависимые каналы составляет 107 ионов в секунду и не может быть меньшей или большей. Канал либо совсем не пропускает вещества (в закрытом состоянии ворот), либо пропускает их с определенной скоростью. Приведенная выше скорость ионного транспорта обеспечивает электропроводность одиночного канала, равную примерно 40 пСм.
Из сказанного следует, что проницаемость мембраны для данного вещества определяется не степенью раскрытия одной поры (она либо сжата, либо полностью раскрыта), а числом открытых каналов в данный момент. Поэтому мембранную проницаемость (р)при переносе веществ по каналам рассчитывают по формуле:
p = 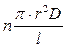 , (47)
, (47)
где п − число открытых каналов на единице поверхности мембраны, r − радиус канала, D − коэффициент диффузии вещества в воде, l − длина канала (примерно равна толщине мембраны).
Переход канала из закрытого состояния в открытое и обратно осуществляется под действием определенных стимулов. Если таким стимулом служит сдвиг мембранного потенциала, то канал относится к группе потенциалзависимых ионных каналов. В них важным компонентом воротного механизма является сенсор напряжения, образованный связанными зарядами, которыми обладают аминокислотные остатки белкового комплекса, составляющего канал. Например, в потенциалзависимом натриевом канале сенсор напряжения, как полагают, образован катионными аминокислотными остатками (гистидилом, лизилом, аргинилом).
Вторую группу каналов составляют потенциалнезависимые каналы. Они управляются не сдвигами мембранного потенциала, а стимулами других модальностей: химическими, механическими, световыми и др. Потенциалнезависимые каналы обычно функционируют во взаимодействии с мембранными рецепторами, воспринимающими соответствующие стимулы. Посредниками между рецепторами и каналами служат, как правило, внутриклеточные сигнальные системы. Подразделение всех каналов на эти две группы − не единственная их классификация. Существуют классификации и по другим признакам, но приведенная классификация очень актуальна, поскольку с функционированием потенциалзависимых ионных каналов связано важнейшее свойство живых тканей − возбудимость.
Кинетика и энергетическое обеспечение трансмембранного переноса гидрофильных веществ через каналы и посредством переносчиков существенно различны. Так, скорость транспорта ионов по каналам, как уже говорилось, составляет 107 ионов в секунду, а посредством переносчиков − 104 ионов · с-1, т. е. в тысячу раз меньше. Потенциальный барьер канального массопереноса − менее 13 кДж · моль-1, а транспорта с участием переносчика − более 67 кДж · моль-1. Коэффициент Бант-Гоффа (Q10), равный соотношению скоростей процесса при температурах, отличающихся на 10о ( 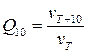 ) , при движении веществ по каналам составляет чуть больше единицы, тогда как переносчик обеспечивает транспорт с Q.10 более 2.
) , при движении веществ по каналам составляет чуть больше единицы, тогда как переносчик обеспечивает транспорт с Q.10 более 2.
По каналам мембрану преодолевают главным образом ионы, а переносчики транспортируют как ионы, так и моносахариды и аминокислоты. Нередко переносчики образуют комплексы с моносахаридом и ионом натрия, с аминокислотой и ионом натрия.
Многие вещества могут транспортироваться разными способами (например, для ионов натрия есть и переносчики, и каналы, работающие одновременно и согласованно). Многообразие вариантов переноса является основой селективной (избирательной) проницаемости разных БМ по отношению к тем или иным веществам. Она зависит от того, какие механизмы и в какой степени сформированы в данной мембране. Так, в одних БМ нет переносчиков моносахаридов, а в других они есть, причем в разных мембранах − в неодинаковых количествах; в одних БМ присутствуют только потенциалнезависимые ионные каналы, а в других − как потенциалнезависимые, так и потенциалзависимые.
В целом при пассивном транспорте селективность БМ определяется коэффициентом распределения вещества между липидом и водой, а в еще большей степени − избирательностью каналов и переносчиков по отношению к определенным агентам. Вместе с тем избирательная проницаемость клеточных мембран тесно связана с работой так называемых биологических насосов.
| <== предыдущая лекция | | | следующая лекция ==> |
| Блочная структура кристаллов | | | Управляемые выпрямители |
Дата добавления: 2015-08-14; просмотров: 7849;