Схема фиксации лидерной последовательности И-РНК на комплементарной последовательности 16S РНК рибосомы
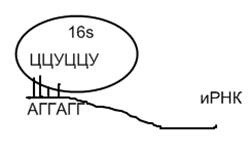
IK – инициирующий кодон АТГ, который кодирует на РНК – АУГ, с него при трансляции начинается синтез полипептида. АУГ в составе молекулы и-РНК на рибосомах кодирует первую аминокислоту белка формилметионин ( у эукариот – метионин). У некоторых микроорганизмов вместо АУГ может быть ГУГ, кодирующий валин.
Зона структурных генов(1-3)начинается сразу за инициирующим кодоном и содержит нуклеотидные последовательности генов, кодирующих ферменты катаболизма лактозы. Для того, чтобы белковые продукты этих генов в процессе трансляции на рибосомах могли разделяться на отдельные белковые молекулы ферментов, между структурными генами моут быть расположены дополнительные последовательности Шайна-Дальгарно.
Терминатор (терминаторный участок)начинается за зоной структурных генов. Он обеспечивает окончание процессов транскрипции (синтеза и-РНК) и трансляции на рибосомах. Терминатор состоит из трех важных блоков: ТК (терминирующего кодона), PD(ГЦ-палиндрома) и ТА-зоны. Рассмотрим их.
ТК -- терминирующий кодонважендля окончания трансляции на рибосомах.Может быть представлен такими триплетами на ДНК: ТАА, ТАГ или ТГА. На и-РНК – это будут стоп-кодоны или нонсенс-кодоны (бессмысленные) УАА, УАГ или УГА. Они не кодируют ни одной аминокислоты, поэтому при трансляции на рибосомах на них обрывается синтез полипептида.
PD -- палиндромы – зона терминатора. Это - инвертированные (повёрнутые на 180 градусов) последовательности, преимущественно из ГЦ- и ЦГ-пар, которые на ДНК способны образовывать крестовую, а на РНК – шпилечную структуру. Для палиндромов характерна комплементарность по горизонтали и вертикали. Именно ГЦ-палиндром в составе терминатора гена (оперона) приводит к замедлению продвижения по данному участку РНК-полимеразы, ведущей синтез и-РНК, т.е. тормозит транскрипцию.
ТА – зона терминатора – участок ДНК, в котором многократно повторяются ТА-пары нуклеотидов, т.е. в смысловой цепи ДНК повторяются остатки тимина, а в матричной - аденина. На матричной цепи ТА-участка ДНК при транскрипции РНК-полимераза синтезирует концевой участок информационной РНК – так называемый уридиловый хвост, состоящий из остатков урацила. Между адениловыми остатками ДНК-овой матрицы и комплементарными уридиловыми остатками синтезируемой и-РНК имеется только по две водородных связи, в отличие от ГЦ-пар, соединенных тремя водородными связями. Поэтому в ТА-зоне происходит легкое отсоединение и-РНК от ДНК-матрицы. Обычно ТА-зона терминатора достаточно протяженная в ρ-независимых (ро-независимых) терминаторах. В ρ-зависимых терминаторах основную роль в терминации (окончании) транскрипции выполняет ρ-белок. Этот белок «садится» на 5/-конец и-РНК, скользит по ней, догоняя РНК-полимеразу и после прокатки через ГЦ- палиндром, сбивает РНК-полимеразу с матричной цепи ДНК. Поэтому ТА зона в ρ-зависимых терминаторах может отсутствовать или быть слабо выражена.
Экспрессия генов у прокариот включает транскрипцию и трансляцию, однако из-за отсутствия ядерной мембраны эти процессы сопряжены и проходят почти одновременно. Освобождающаяся от матричной ДНК молекула и-РНК поступает на полисому (комплекс ≈ из 10 рибосом), которая располагается на ЦПМ, где почти сразу начинается трансляция. Белковый продукт появляется в клетке через 2,5-3 минуты после начала транскрипции.
Регуляция экспрессии генов осуществляется на уровне транскрипции и трансляции. На уровне транскрипции с помощью таких механизмов, как негативный и позитивный контроль, индукция и репрессия, аутогенный контроль, катаболитная репрессия. На уровне трансляции изучен механизм регулирования с помощью образования альтернативных шпилек на и-РНК - аттенуация.
In vivo (в живой клетке) чаще всего имеют место смешанные механизмы регулирования. Выделяют 4 типа классических оперонов:
1) индуцибельный с положительным контролем;
2) индуцибельный с отрицательным контролем;
3) репрессибельный с положительным контролем;
4) репрессибельный с отрицательным контролем;
Встречаются и смешанные механизмы регуляции, например, катаболитная репрессия.
Передача генетической информации у прокариот осуществляется как по вертикали (от материнской клетке к дочерней), так и по горизонтали (между клетками одного вида, разных видов, родов, семейств) с помощью механизмов рекомбинации – конъюгации, трансформации, трансдукции, сексдукции, трансфекции, транспозиции. Горизонтально, могут передаваться плазмиды, транспозоны, вирусы. Все они могут захватывать и переносить фрагменты генома.
Для прокариотов, кроме хромосомного, характерен еще плазмидный геном.В клетке может находиться от нескольких единиц до нескольких сотен плазмид.
Плазмиды – двухцепочечные кольцевые ДНК, которые несут дополнительные гены: гены устойчивости к токсинам, антибиотикам, сульфаниламидам, тяжёлым металлам, нефтепродуктам, нафталину, камфоре, а также гены синтеза токсинов, антибиотиков, гемолизинов и других факторов патогенности. Гены факторов патогенности обычно собраны в особые кластеры (группы), получившие название островов патогенности. С помощью плазмид, фагов и транспозонов острова патогенности и другие геномные острова (Genom islands) могут переноситься горизонтально от одной клетки к другой не только в пределах вида, но и между представителями разных родов и семейств.
В 70-х годах у бактерий обнаружили транспозоны – мобильные генетические элементы (так называемые «прыгающие» гены), способные перемещаться с одного сайта генома в другой, например, между различными участками хромосом, из хромосомы в плазмиду, в фаговую ДНК и наоборот.
Дата добавления: 2015-08-08; просмотров: 1660;