Кардиомиоциты
Сравнительная гистология сердечной мышечной ткани
Структура сердечной мышечной ткани животных.
Кардиомиоциты
Сердечная мышца позвоночных имеет клеточное строение и представлена тремя субпопуляциями кардиомиоцитов:
 1 - кардиомиоциты желудочков
1 - кардиомиоциты желудочков
2 - кардиомиоциты предсердий «рабочие миоциты»
3 - миоциты проводящей системы сердца
Структура миоцитов сердца детально изучена в работах П.П.Румянцева и его учеников (70-90 е годы ХХ века).(Румянцев, 1990).
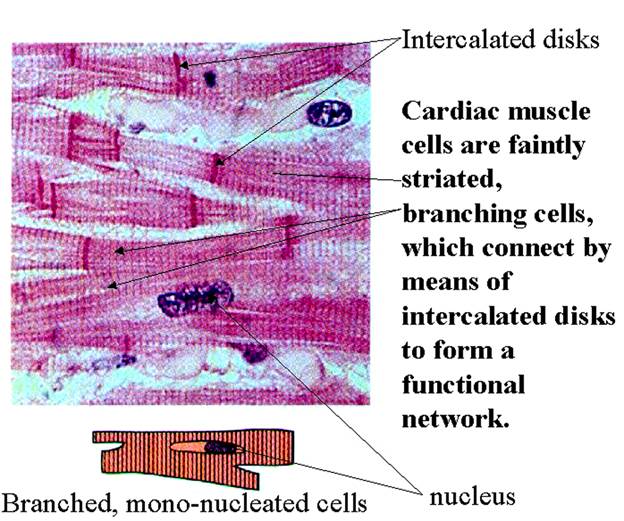
Рис. 4.1.1. Микрофотография и схема строении кардиомиоцитов
Отдельная клетка миокарда сердца имеет довольно крупные размеры (табл. 1), как правило, одно- два крупных ядрер и хорошоразвитую систему миофибрилл.
У человека доля дву- ядерных миоцитов составляет в желудочке 10-13%, а многоядерные клетки составляют меннее 1%.
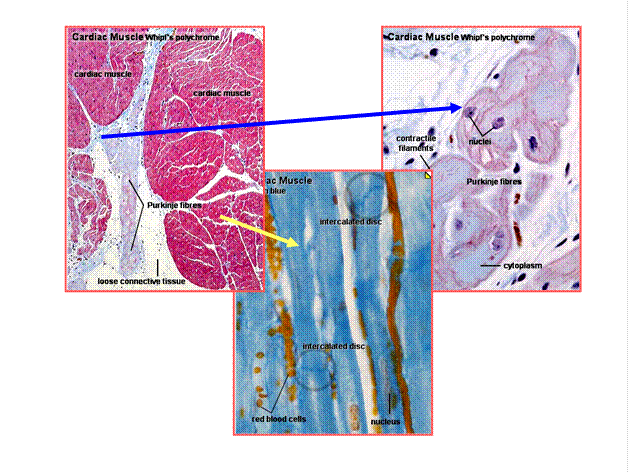
Рис. 4.1.2. Гистология сердечной мышцы млекопитающих (кардиомиоциты и клетки волокон Пуркинье)
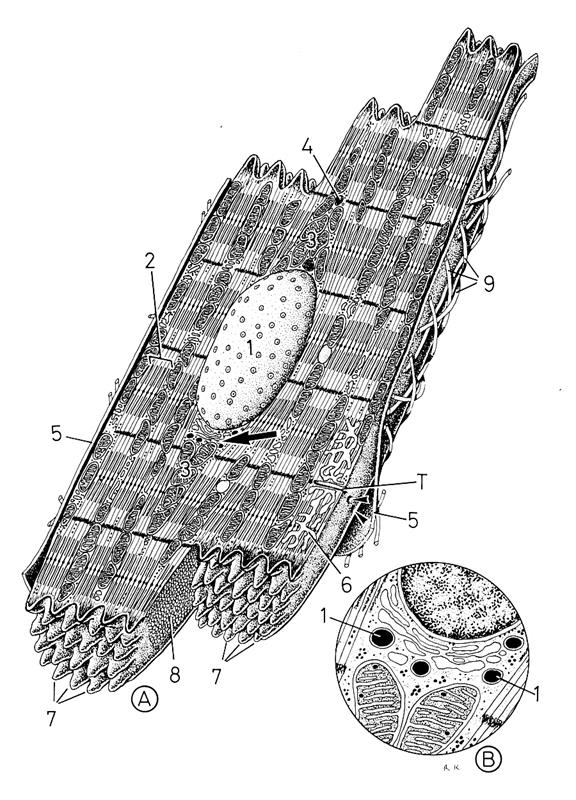
Рис 4.1.3. Ультраструктура кардиомиоцита желудочков сердечной мышцы млекопитающих (по: Крстич, 2001)
Среди особенностейорганизации кардиомиоцитов (по сравнению со скелетными мышцами) следует отметить следующие:
- сильное развитие митохондрий особого «цепочечного типа». Отдельные митохондрии располагаются между миофибриллами,образуя длинные цепочки. Каждая митохондрия соединена в соседней с помощью специализированного межмитохондриального контакта (ММК).Они образуют единую энергетическую систему миоцита и есть в сердечной ткани всех позвоночных, а также у моллюсков и насекомых. Кроме выше описанных т.н. «цепочечных, межфибриллярных»митохондрий, в миоцитах есть еще две субпопуляции митохондрий: субсарколеммальная – группы митохондрий, расположенных в местах прилежания капилляров и околоядерная - группы митохондрий концентрирующиеся в ядерной области. Во всех субпопуляциях между митохондриями есть ММК.
-L – система в кардиомиоцитов развита слабее, чем в скелетных мышцах, образуя сплошную сеть мелких продольных и поперечных анастомазирующих канальцев и цистерн. В отличие от скелетных мышц, каналы L –системы образуют не только триады и диады в месте контакта с Т-трубочками и сарколеммой, но контактируют с сарколеммой в различных участках миоцита, образуя т.н. субсарколеммальные цистерны.Общий объем, занимаемый L-системой в кардиомиоците составляет около 4% его объема.
- Т-система, наоборот, развита значительно сильнее, чем в скелетной мышце. Т-каналы имеют средний диаметр в 150-200 мкм (против 40 мкм в скелетных мышцах) и имеют вид широких разветвленных каналов, входящих в кардиомиоцит на уровне z – дисков. Полагают, что по Т-системе в сердце распространяется не только импульс, но и поступают в клетку метаболиты.
Объем Т-системы составляет до 35% объема клетки ( в скелетных мышцах – 0,5%).
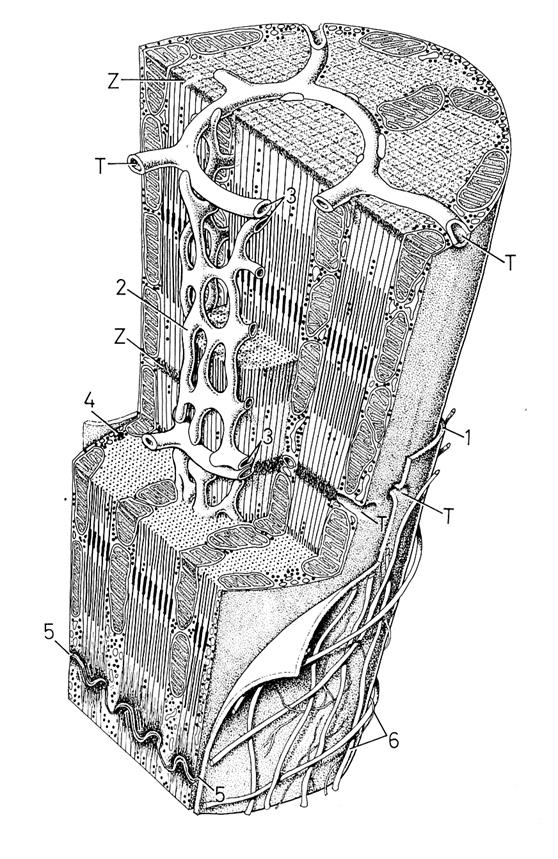
Рис. 4.1.4. Схема ультраструктурной организации кардиомиоцита млекопитающих
(по: Крстич, 2001)
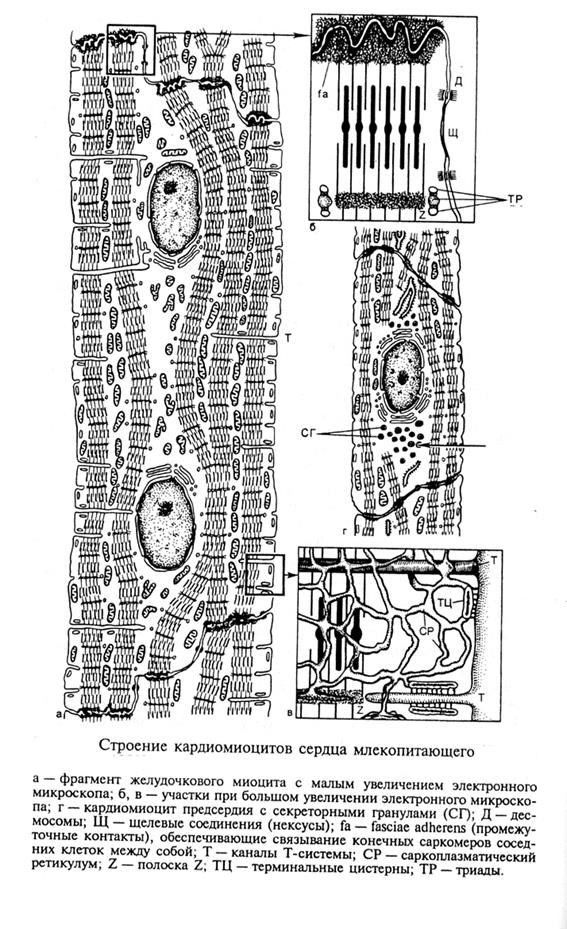
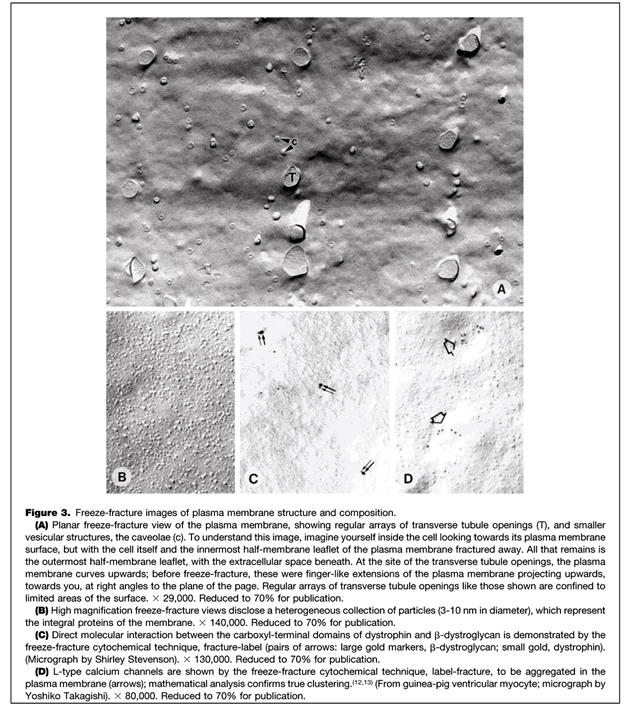
Рис. 4.1.5. Фрагменты организации Т-системы кардиомиоцитов млекопитающих
(по: N.Severs, 2000)

Рис. 4.1.6. Фрагменты организации L-системы в кардиомиоцитах млекопитающих
(по N.Severs, 2000)
- присутствие в сердкечной мышцы специальных межклеточных контактов между кардиомиоцитами - вставочных дисков (intercalated discs). Это специализированная структура сердечной мышцы, предназначенная для соединения отдельных миоцитов в единую морфо-функциональную системы мышечных пучков (волокон) миокарда.
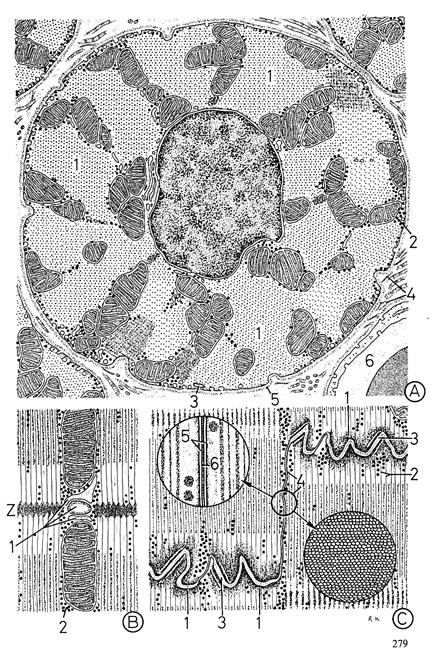
Рис. 4.1.7. Элементы структуры кардиомиоцита млекопитающих
(по; Крстич, 2001)
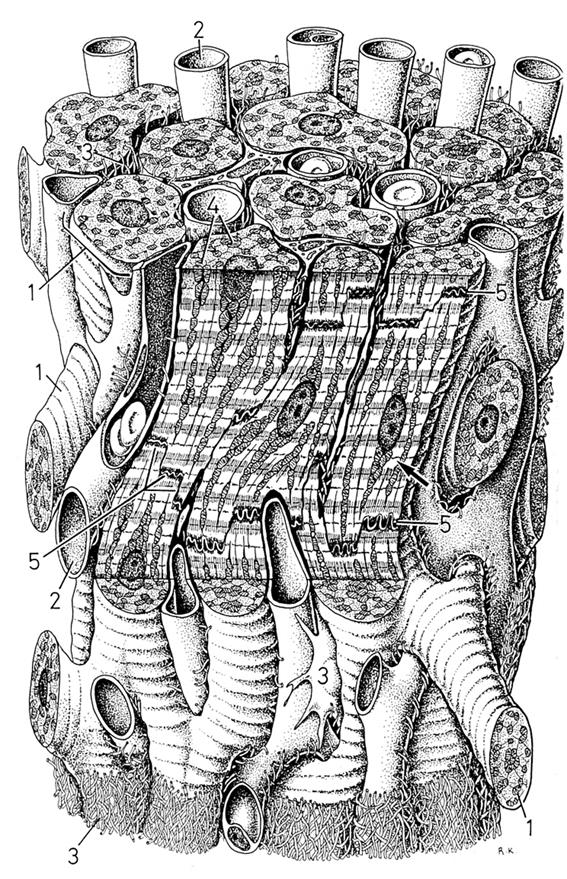
Рис. 4.1.8. Схема строения пучка кардиомиоцитов в миокарде млекопитающих.
Хорошо видна система вставочных лисков (5), соединяющих отдельные клетки в пучок. (по: Крстич, 2001)
В состав ИД входит комплекс специализированных контактов: промежуточные соединения ( fascia adherens), щелевые контакты (gap junction) и десмосомы (desmosomes). В их составе множество специализированных белков, они всегда проходят на уровне Z- дисков и связаны с сарколеммой. (см. Лекцию 1, строение сарколеммы).

Рис.4.1.9. Cтруктура интеркалярных дисков кардиомиоцитов
(по: N.Severs, 2000)
Промежуточные соединения и десмосомы предназначены для соединения миофибрилл с десминовым цитоскелетом и кадхериновыми контактами соседних плазматических мембран миоцитов, а gap junction предназначен для передачи электрических импульсов между миоцитами при возбуждении сокращения.
Коннексоны, входящие в состав щелевых контактов состоят из шести коннексинов- 43, в каждой клетке во вставочном диске имеет целая система дисков. В центре находится канал, способный пропускать молекулы до 1KD ( инозитол фосфат, цАМФ, ионы Са и др).
Не все коннексины одинаковые – однаружено до 16 генов, которые контролируют экспрессию разных видов коннексинов ( коннексин-43, 40,45). Разные коннексины обнаружены в составе контактов в разных участках миокарда, по-видимому определяя какие-то функциональные различия этих участков миокарда. Располагаются они вдоль длинной оси миоцитов, тогда как десмосомы и промежуточные соединения (зоны слияния) поперек, и обладают низким электрическим сопротивлением.
- миоциты предсердий помимо сократительной функции, обладают секреторной активностью, синтезируя ряд гормонов:
1. - кардиодилатин и его производное - кардионатрин (предсердный натрийуретический пептид),
(он вызывает сокращение гладких мышц артерий, увеличивает почечный кровоток, ускоряет фильтрацию и выделение натрия)
2 -энкефалининвертаза – (пептид, регулирующий синтез энкефалина и других «опиоидных» нейропептидов
- кардиомиоцитыв большей степени, чем скелетные мышцы, зависят от концентрации ионов Са во внеклеточной жикости. Именно Са из внеклеточной жидкости попадая в цитоплазму кардиомиоцитов индуцирует Са-зависимый мехенизм освобождения Са из цистерн гладкого ретикулума. Изменение количества ионов Са, высвобождающегося из цистерн ретикулума сильнее влияет на сокращение сердечной мышцы, чем Са в L - цистернах скелетной мышцы. В связи с этим свойством, любое изменение концентрации ионов Са во внеклеточной жидкости и изменение Са проницаемости кардиомиоцитов (медекаментозное, гормональное, вегетативное) очень сильно влияет на работу сердечной мышцы.
- предсердные миоциты, согласно таблице, приведенной выше,мельче желудочковых миоцитов и имеют ряд специфических черт строения: несколько ядер, иногда полиплоидные (до 8с), хорошо развитая система ШЭC и АГ, система секреторных гранул.
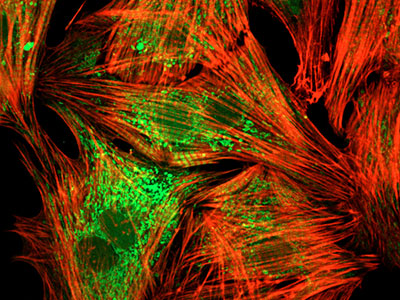
Рис. 4.1.10. Иммуноцитохимия кардиомиоцитов (в культуре). Красным обозначены фибриллы актинового цитскелета, зеленым – бактериальная инфекция (эндотоксины) при сепсисе.
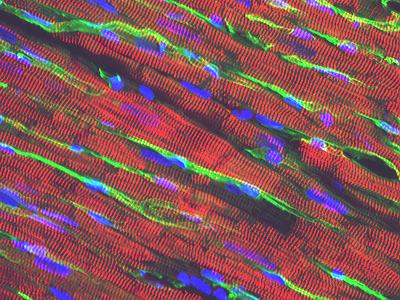
Рис. 4.1.11 Иммуногистохимия кардиомиоцитов. Красный цвет – а-актинин, зеленый – кровеносные сосуды, голубой – ядра кардиомиоцитов и других клеток.
4.2. Клетки проводящей системы сердца млекопитающих.К проводящей системе сердца относят группу узлов (синусно-предсердный, предсердно-желулочковый и проводящие пучки, пронизывающие весь миокард)
Клетки синусно-предсердного узла являются самыми главными в работе проводящей системы и обладают ритмической (пейсмекерной) активностью. Они довольно сильно отличаются по морфологии от обычных кардиомиоцитов:
- имеют мелкие размеры (20-30 х 8 мкм)
- очень мало миофибрилл и митохондрий
- довольно бедная органоидами цитоплазма
- состоят из двух субпопулляций – центральная группа мелких клеток (Р клетки)и периферические более крупные клетки (с большими числом миофибрилл). последние передают импульсы в пучки проводящей системы сердца.
Клетки предсердно-желудочкового узлавесьма гетерогенны по размерам, но по своей морфологии напоминают клетки СПУ: они также бедны миофиламентами, имеют слабо развитую систему вставочных дисков и системы L и Т- каналов
Клетки проводящих пучков ( гисса, Кента и др) и клетки Пуркиньенаряду с
общими чертами, имеют четко выраженные систематические различия. Условно выделяют три группы млекопитающих с разным типом строения клеток Пуркинье и пучков
1 – крупные млекопитающие и, в первую очередь, копытные (овцы, козы, лошади, коровы, свиньи). У этих млекопитающих клетки Пуркинье самые крупные (диаметром до 80-100 мкм), но с малочисленными, периферически расположенными миофибриллами и широкой центральной частью, занятой саркоплазмой с несколькими ядрами, аппаратом Гольджи и многочисленными гранулами гликогена.
2 – млекопитающие средних размеров(собаки, кошки) и человек. У этих млекопитающих клетки Пуркинье более мелкие и их популяция более гетерогенна, чем в первой группе. Так, если в левой ножке Гиса преобладают типичные клетки Пуркинье со слаборазвитой системой миофибрилл, то в правой, напротив, клетки содержат много пучков миофибрилл и напоминают по структуре типичные кардиомиоциты. Среди них вcnречаются и гигантские клетки, напоминающие клетки Пуркинье копытных.
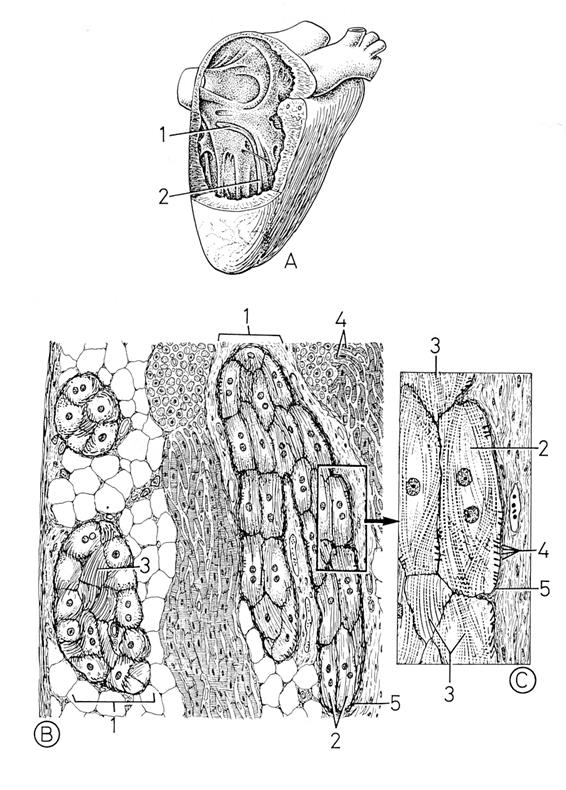
Рис. 4.2.1. Строение элементов проводящей системы сердца. Световой микроскоп
(по Крстич, 2001)
3 – мелкие млекопитающие (мыши, крысы, кролик, хорек и др). У этих млекопитающих типичные клетки Пуркинье встречаются только в составе ножек Гиса и в районе их разветвления в желудочках. В волокнах Пуркинье клетки проводящей системы все меньше и меньше отличаются от соседних рабочих кардиомиоцитов – они имеют хорошо развитую систему миофибрилл, четкие вставочные диски, систему L-каналов и терминальных цистерн (хотя по общему количеству миофибрилл они все-же уступают кардиомиоцитам:
хорек– клетки пучка Гиса объем миофибрилл - 27-33%
предсердно-желудочковый узел - 32%
рабочие кардиомиоциты - 54%
Морфологические различия рабочих и проводящих миоцитов сердца сопряжены со значительными различиями в цитохимии этих клеток. В клетках проводящей системы обнаружено:
- значительное количество гликогена
- низкая активность основных ферментов цикла Кребса и дыхательной цепи ( сукцинатдегидрогеназы, изоцитродегидрогеназы, цитохромоксидахы и др)
- более активны ферменты гликолизного цикла ( малатдегидрогеназа, лактатдегидрогеназа), что свидетельствует о преобладании анаэробных процессов в клетках проводящей системы
- высокая активность ферментов лизосомальной системы
Клетки проводящей системы имеют богатую систему иннервации, в основном адренэргические и катехоаминэргические эфферентные окончания и многочисленные афферентные окончания.


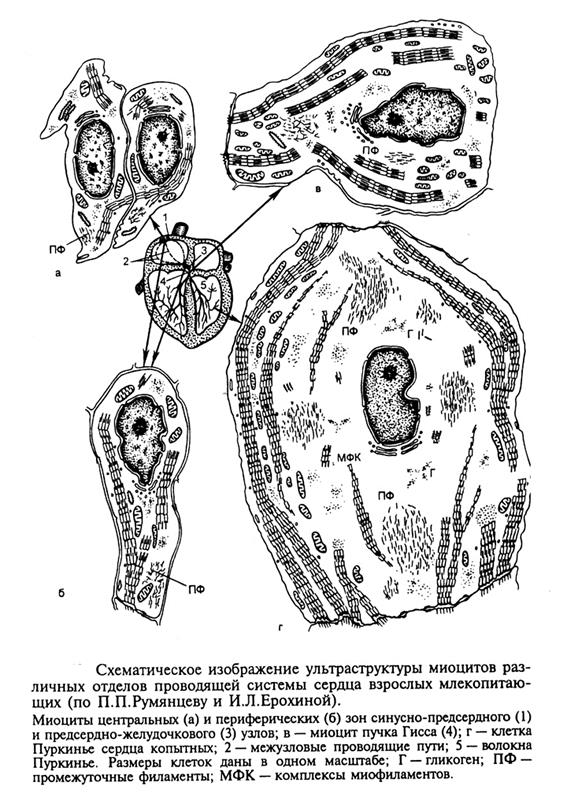
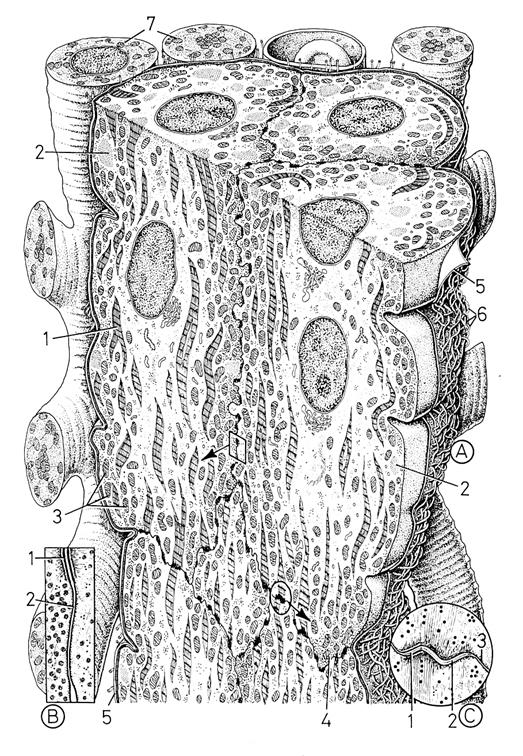
Рис. 4.2.3. Схема ультраструктуры клеток волокон Пуркинье
(по Крстич, 2001, Шубникова и др, 2001)
Дата добавления: 2015-07-07; просмотров: 11739;