Строение и состав миофибрилл сердечной мышечной ткани позвоночных. Строение и особенности саркомеров миофибрилл сердца.
В принципе, структура саркомера в миофибриллах кардиомиоцитов не отличается от саркомеров скелетной мышцы, но есть ряд биохимических отличий, связанных в большей степени с особенностями регуляции процесса сокращения и строением самих мышечных клеток сердца.
- особые изоформы тяжелых и легких цепей миозина. В сердце присутсвует две формы тяжелых цепей и четыре формы легких цепей миозина, которые теоретически могут сформировать множество разновидностей миозиновых молекул. На практике в желудочках обнаружено три формы V1,2,3 и две в предсердиях A1,2.
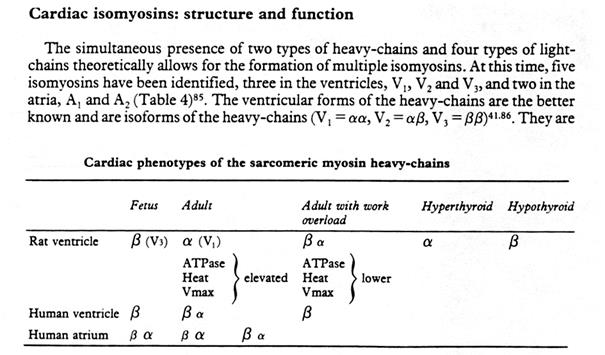
Распределение типов миозинов также зависит от вида и стадии эмбрионального развития
у крысы, кролика,собаки, коровы, человека в эмбриональный период и после рождения – в желудочке β-МНС и α – МНС в предсердиях к моменту созревания у всех видов в желудочках появляется α – МНС форма и β-МНС в предсердиях. В конце концов формируется определенная мозаика распределения изоформ миозина и его цепей в желудочках и предсердиях

- Особые изоформы и характер распределения изоформ актина, также зависящие от возраста и вида животного.
В целом имеется две формы актина – скелетный (a-sk) и сердечный (a-card), которые структурно очень похожи, но различаются только по четырем АК в позиции 375. Обнаружены также различия в размере и РНК актинов – в сердечной и_РНК 5” конец молекулы на девять нуклеотидов длиннее, чем в скелетной и-РНК..

- в саркомерах сердца имеются свои формы титина: N2BA и NB2, которые отличаются от титина скелетных мышц по молекулярному весу и по видовому признаку. У крупных млекопитающих есть обе формы титина, у кроликов – только NB-2 форма. Титин, как это отмечалось ранее, играет важную роль в сокращении и расслаблении мышцы, во многом определяя взаимозависимость степени растяжения мышцы и силы ее сокращения. Возможно именно свойствами титина и определяются существенные различия в этих соотношениях между сердечной и скелетной мышцей.
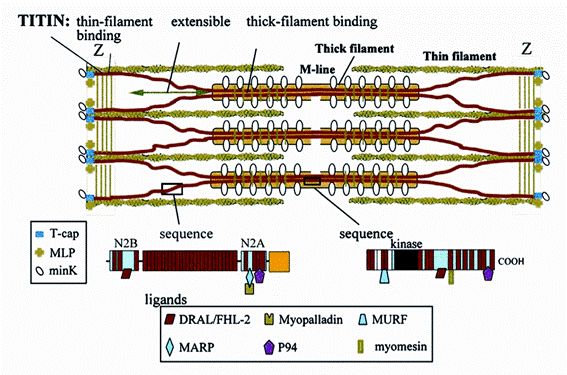
Рис. 4.3.1. Схема строения саркомера кардиомиоцита млекопитающих
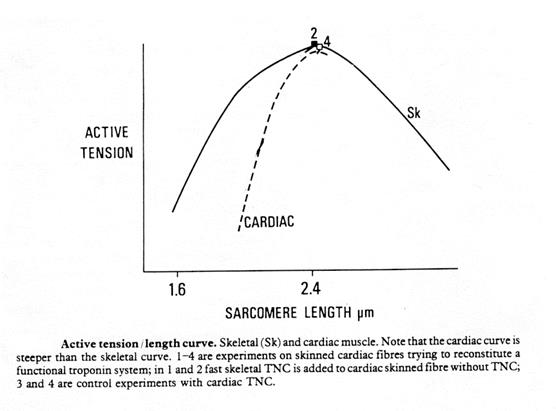
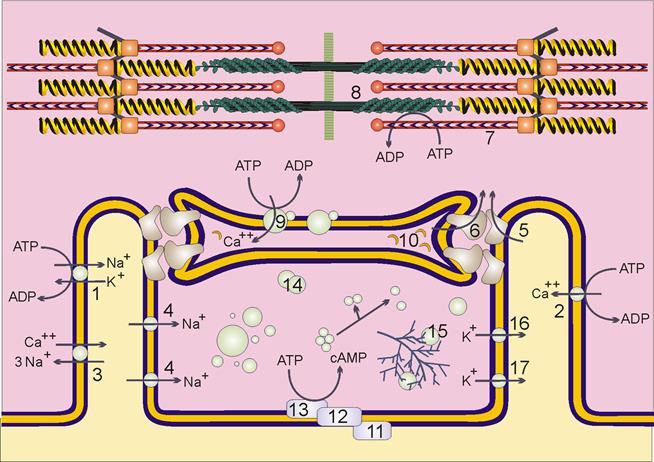
Рис. 4.3.2. Схема саркомера сердечной мышцы и ионных потоков, участвующих в сокращении
The diagram above shows the control of cardiac muscle contraction. The Na/K ATPase or sodium pump (1) works continuously, using the energy from ATP to maintain a high K concentration inside the cells and a high Na concentration in the extracellular fluid (ECF). The cell membrane (sarcolemma) is usually more permeable to potassium ions than to sodium ions, and this gives rise to a membrane potential of about 80mV (inside negative) in relaxed muscle. Calcium ions are also removed from the cytosol into the ECF by an ATP-driven calcium pump (2) in all tissues. Cardiac muscle possesses an additional sodium/calcium exchange protein (3). This export system is driven by the pre-existing sodium ion gradient. The calcium concentration inside resting cells is low (0.01mM) , but rises sharply to 1 mM during contractions. The sarcolemma is very thin (about 6 nm) so the 80mV membrane potential equates to a voltage gradient of about 13,000,000 volts per metre! All membrane components are subject to intense electric fields, and protein conformations are greatly influenced by the membrane potential. "Voltage gated" ion channels will only conduct over a narrow range of membrane potentials, whereas "ligand gated" ion channels (such as the acetylcholine receptor in voluntary muscle) require specific chemical activators.
Muscle contraction is triggered by a wave of membrane depolarisation which spreads from neighbouring cardiac cells. [In skeletal muscle the cells work independently and are triggered by their motor neurone via the motor end plate.] The change in electric field activates voltage gated sodium channels (4) in the sarcolemma, each of which allows a few hundred positively charged sodium ions to enter the negatively charged cytosol, further reducing the membrane potential until the whole sarcolemma is depolarised. These sodium gates close spontaneously after a few milliseconds, but slower voltage-gated calcium ion channels, thought to be identical with the dihydropyridine receptors (5) take over and maintain a positive inward current for several hundred milliseconds (in human ventricle) during the plateau phase of the cardiac action potential. [The plateau phase is absent in skeletal muscle.] The membrane potential is eventually restored to its resting value by a delayed loss of positive potassium ions (16) from the cells.About 10% of the calcium needed to activate cardiac contraction enters during each beat from the ECF. The remainder is released from the sarcoplasmic reticulum through a channel known as the ryanodine receptor (6). Separate genes coding for this enormous protein (over 5000 amino acids) are active in heart and skeletal muscles. Operation of the ryanodine receptor depends in a mysterious way on the flow of calcium ions through the dihydropyridine receptors in cardiac muscle, but not in skeletal muscle.
Таким образом, видно, что сократимость сердца в значительной степени зависит от притока ионов Са извне. Любые стимулы, которые могут приводить к продолжительному увеличению концентрации Са в цитоплазме (за счет повышенного притока Са из межклеточного пространства, освобождение Са из цистерн ЭР, или пониженного выхода Са из цитоплазмы) вызывают увеличение сократимости.
Например: катехоламины, действуя через метаботропные в-адренорецепторы, через цАМФ фосфорилируют Ca-L каналы, что приводит к увеличению притока Са извне в кардиомиоцит, а повышение концентрации Са в цитоплазме активирует Са каналы в саркоплазматическом ретикулуме, вызывая дополнительный приток Са в район миофибрилл.
Сердечные гликозиды (например дигиталис) подавляют Na,K –насос, что вызывает уменьшение мембранного градиента Na и уменьшает выход Са из цитоплазмы через Na, Ca – насос в плазматической мембране клетки. Стимуляция Вагуса, напротив приводит к подавлению сократимости, вследствие подавления притока ионов Са в клетку.
В отличии от скелетных мышц сердечная мышца обычно работает при намного меньших растяжениях, чем оптимальная длина волокна.
Дата добавления: 2015-07-07; просмотров: 2758;