НАСЛЕДУЕМОСТЬ ПОКАЗАТЕЛЕЙ КОЖНО-ГАЛЬВАНИЧЕСКОЙ РЕАКЦИИ
Изучение кожно-гальванической реакции (КГР) впервые началось в конце XIX в., когда почти одновременно французский невропатолог К.Форе и российский физиолог И.Р. Тарханов зарегистрировали; первый — изменение сопротивления кожи при пропускании через нее слабого тока, второй — разность потенциалов между разными участками кожи.
Электрическая активность кожи обусловлена главным образом активностью так называемых эккринных потовых желез в коже человека, которые в свою очередь находятся под контролем симпатической нервной системы. Главная функция этих желез —терморегуляция, т.е. поддержание постоянной температуры тела, Однако эккринные железы, расположенные на ладонях и подошвах ног, а также на лбу и под мышками, реагируют в основном на экстренные внешние раздражители и стрессогенные воздействия, поэтому КГР, как правило, регистрируют с кончиков пальцев или ладоней.
В психофизиологии электрическую активность кожи используют как показатель «эмоционального» потоотделения. КГР возникает также в ответ на изменения во внешней среде (как компонент ориентировочной реакции) и имеет большую амплитуду при большей неожиданности, значимости и интенсивности стимула. При повторных предъявлениях стимула КГР постепенно снижается, этот процесс называется привыканием. Следует, однако, иметь в виду, что подлинная природа КГР до сих пор неясна,
Амплитуда и скорость привыкания КГР имеют высокую межиндивидуальную вариативность и при соблюдении постоянства условий регистрации обнаруживают относительно высокую воспроизводимость, что позволяет исследовать роль генотипа в межиндивидуальной дисперсии данных показателей. Первые исследования роли факторов генотипа в происхождении индивидуальных особенностей КГР проводились в 60-70-х годах. Их результаты оказались противоречивыми. Так, С. Ванденберг и его коллеги не выявили достоверных различий МЗ и ДЗ близнецов по амплитуде КГР на разные стимулы [435]. У. Хьюм, изучая наследуемость параметров КГР на звуки и холодовое воздействие, обнаружил умеренный вклад наследственных влияний в изменчивость амплитуды и скорости привыкания КГР на звук 95 дБ, для тех же параметров КГР на холодовое воздействие влияний генотипа установить не удалось [292].
Тем не менее по мере накопления данных становилось все очевиднее, что индивидуальные параметры КГР относятся к числу генетически обусловленных характеристик (табл. 15.1). В целом ряде исследований было установлено: МЗ близнецы по сравнению с другими парами близких родственников имеют более высокое внутрипарное сходство по таким показателям КГР, как амплитуда, латентный период и скорость привыкания, что дало основание говорить о влиянии генотипи-ческих факторов на межиндивидуальную изменчивость и этих показателей, и реакции в целом. МЗ близнецы также более схожи, чем ДЗ, по показателям времени восстановления КГР после воздействия и скорости роста КГР до максимального значения [203].
Наиболее полное генетическое исследование КГР было проведено Д.Ликкеном с соавторами [327]. Параметры КГР на громкие звуковые стимулы исследовались на большой выборке близнецов, часть которых с раннего детства воспитывалась в разных семьях. Анализировались следующие показатели: максимальная амплитуда КГР, средняя амплитуда КГР в первых четырех пробах, показатели снижения амплитуды по мере привыкания и ряд других. При этом учитывались не только абсолютные значения амплитуды КГР, но и относительные, которые определялись как частное от деления амплитуды каждой отдельно взятой реакции данного испытуемого к максимальному значению амплитуды, зафиксированной у него через 3 с после первого предъявления звукового сигнала интенсивностью 110 дБ.
Было обнаружено, что дисперсия параметров КГР в значительной степени обусловлена генотипом (табл. 15.2). Коэффициенты корреляции, характеризующие внутрипарное сходство МЗ близнецов, превышали оценки внутрииндивидуальной стабильности КГР. Воспроизводимость параметров КГР характеризуется коэффициентами корреляции от 0,5 до 0,6. Иначе говоря, сходство КГР у МЗ близнецов оказалось даже выше, чем сходство КГР у одного и того же человека при повторных регистрациях. У ДЗ близнецов аналогичные коэффициенты были значительно ниже. Причем наиболее значительные различия были получены для абсолютных показателей, в этом случае сходство ДЗ близнецов было намного ниже, чем МЗ. Подобная разница позволяет относить абсолютные значения параметров КГР к категории признаков, которые Д. Ликкен назвал эмерджентными. Они определяются не семейным сходством, а уникальными особенностями сочетания генов конкретного генотипа [326].
При использовании относительных значений сходство МЗ близнецов примерно в два раза превышало сходство ДЗ, что свидетельствует об аддитивном действии генов. С точки зрения авторов, различия в характере генетических влияний — эмерджентный для абсолютных и аддитивный для относительных оценок амплитудных параметров КГР — объясняются разной природой данных показателей. При этом подразумевается, что абсолютная амплитуда КГР определяется боль-
Таблица 15.1
| Наследуемость параметров КГР (по данным разных | авторов) | ||||
| Автор, | Испытуемые, возраст | Стимулы | Параметры КГР | Генетический | Основные |
| год публикации | (в годах) | анализ | результаты | ||
| С. Ванденберг и др. | вспышка, | сопротивлениеКОХИ | дисперсионный | КГР не наследуема | |
| (С. Vandenberg et al.), | стук, звонок | анализ | |||
| B.X96ьюм (W.Hume), | звук 95 дБ, | амплитуда, | внутриклассовые | умеренная наследуемость КГР на звук; | |
| 1000 Гц. | скорость привыка- | корреляции | отсутствие наследуемости КГР на хо- | ||
| холодовое | ния КГР | лодовое воздействие | |||
| П. Звольский | 15 пар МЗ | звуки разной | площадь под | внутриклассовые | наследуемость КГР на звук в покое: |
| (P.Zvolsky et al.), 1976 | 19 пар ДЗ | силы, покой, | кривой КГР | корреляции | Н = 0,5 и при испуге: Н = 0,4 |
| 18-1Q | ГТПРГГ | ||||
| Б. Кочубей, 1983 | 22 пары МЗ | звуки 80 дБ, | амплитуда, ла- | дисперсионный | все показатели наследуемы |
| 21 параДЗ | 105 дБ, 1000 Гц | тентный период, | анализ | G составляет для амплитуды 50-63%, | |
| 17-29 | интенсивность ТПЛТТТЗ Т_ТХГ Q ХТТТ а | для латентного периода 82— 73%, для | |||
| Д. Ликкен (D. Lykken | 36 пар МЗ близнецов, | звук 110 дБ | амплитуда, | метод подбора | все показатели наследуемы |
| etal.), 1988 | выросших вместе | интенсивность | моделей | аддитивное наследование для относи- | |
| 43 пары ДЗ | привыкания | тельных показателей, неаддитивное — | |||
| 42 пары разлученных МЗ | (абсолютная, | для абсолютных | |||
| близнецов | относительная) | ||||
| 16-56 | |||||
| Т. Бушар (Т. Bouchard | 17 мужских пар | звук 110 дБ | амплитуда, | внутриклассовые | у мужчин наследуемость амплитуды |
| etal.), 1990 | 19 женских пар МЗ, рос- | интенсивность | корреляции | КГР выше: rМЗ (разлученных) 0,82; | |
| ших вместе | привыкания | у женщин ниже: rМЗ (разлученных) | |||
| 20 мужских пар | 0,30 | ||||
| 23 женские пары разлу- | |||||
| ченных МЗ близнецов | |||||
| 16-56 |
Таблица 15.2 Наследуемость параметров КГР [по данным 327]
| Показатели | Коэффициенты корреляции | Доли вариативности | ||||
| МЗДМ(36 пар] | МЗР(43 пары) | ДЗ (42 пары) | генетическая | средовая | ||
| общая | индивидуальная | |||||
| Абсолютные :крутизна наклона кривой, характеризующей привыкание КГР число предъявлений стимула до исчезновения ответа средняя амплитуда КГР из первых четырех проб максимальная амплитуда КГР | 0,72 0,42 0,62 0,45 | 0,54 0,43 0,52 0,37 | 0,05 0,13 0,10 0,13 | 70,3 40,7 59,1 56,0 | 5,2 00 00 | 24,5 58,3 40,9 44,0 |
| Относительные :относит ельная амплитуда КГР (отношение средней к максимальной) | 0,66 | 0.58 | 0,35 | 54,1 | ид | 34,8 |
Примечание. МЗВ — монозиготные близнецы, воспитанные вместе; МЗР — разлученные монозиготные близнецы.
шим числом факторов и отражает не только реактивность ЦНС на внешнее воздействие, но и некоторые побочные эффекты (например, она зависит от числа потовых желез в месте приложения электрода и др.). При вычислении относительных величин посторонние влияния исключаются, что и приводит к изменению соотношения компонентов наследуемости.
Вместе с тем, по данным Д. Ликкена, в изменчивость параметров КГР существенный вклад вносит и индивидуальная среда (табл. 15.2), что, по-видимому, неслучайно, поскольку динамика КГР тесно связана с ориентировочно-исследовательской активностью индивида и особенностями его эмоционального реагирования, которые в онтогенезе претерпевают существенные изменения.
2. наследуемость показателей функционирования сердечно-сосудистой системы
Показатели работы сердечно-сосудистой системы используются в психофизиологии как источник информации об изменениях, происходящих в организме в связи с различными психическими процессами и состояниями.
Индикаторы активности сердечно-сосудистой системы включают: частоту сердечных сокращений (ЧСС); силу сокращений сердца, т.е. силу, с которой сердце накачивает кровь; минутный объем сердца — количество крови, проталкиваемое сердцем в одну минуту; артериальное давление (АД); региональный кровоток — показатели локального распределения крови.
В целом ряде исследований, выполненных на близнецах, было показано влияние генотипа на индивидуальные особенности ЧСС, а также давления крови в состоянии покоя и при различных нагрузках (табл. 15.3).
По данным разных авторов, оценки наследуемости ЧСС и кровяного давления варьируют в широких пределах: для показателей ЧСС от 0 до 70%, для показателей давления от 13 до 82% (систолического) и от 0 до 64% (диастолического), составляя в среднем 50% [84, 141, 200, 201], Степень наследственной обусловленности показателей кровяного давления, по-видимому, может изменяться с возрастом: отмечается тенденция к снижению влияния наследственных факторов на уровень диастолического давления у взрослых при переходе от молодого к зрелому возрасту (от 68 к 38%), что происходит за счет увеличения влияния несистематической среды. Вероятно, существуют также половые различия в наследуемости показателей систолического и диастолического давления, но однозначно определить характер этой зависимости пока не представляется возможным.
Ввиду того, что показатели работы сердечно-сосудистой системы существенно и закономерно изменяются в условиях деятельности, особый интерес представляют исследования природы межиндивидуальной дисперсии реактивных изменений ЧСС, давления и других показателей такого рода, точнее их изменений, которые возникают в ходе выполнения различных задач.
Одно из наиболее развернутых исследований в этом плане было проведено Е.И. Соколовым с соавторами [141]. У 24 пар МЗ близнецов и 19 пар ДЗ они регистрировали показатели давления (систолического, диастолического и общего), ЧСС, а также показатели кровенаполнения сосудов головного мозга (реографический индекс). Перечисленные показатели регистрировались в трех экспериментальных ситуациях: покое, при психоэмоциональной нагрузке и через 10 мин после нее. Нагрузка — интеллектуальная деятельность в условиях дефицита времени с действием отвлекающего раздражителя (свет, звук).
22-1432
Таблица 15.3
Наследуемость показателей работысердечно-сосудистой системы (по данным разныхавторов)
| Автор, год публикации | Испытуемые, возраст (в годах) | Условия | Показатели | Генетический анализ | Основные результаты |
| Дж. Мазер и др. (J. Mather et al.), 1961 | 34 пары МЗ 19 пар ДЗ 29 | Покой | QRS -QT— интервалы в ЭКГ, ЧСС | дисперсионный анализ | по QRS-QT— интервалам МЗ более похожи, чем ДЗ; по ЧСС сходство одинаково |
| С Ванденберг (С. Vandenberg et al.), 1965 | вспышка, стук, звонок | ЧСС | дисперсионный анализ | МЗ более похожи, чем ДЗ | |
| А. Шапиро и др. (A. Shapiro et al.), 1968 | 12 пар МЗ 12 пар ДЗ | тест Струпа, болевые стимулы | давление, ЧСС | дисперсионный анализ | МЗ более похожи, чем ДЗ |
| В. Хьюм (W. Hume), 1973 | покой, звук 95 дБ, 1000 | ЧСС | внутриклассовые корреляции | отсутствие наследуемости ЧСС | |
| В. Клисорас и др. (V. Klissouras et al.), 1973 | 23 пары МЗ 16 пар ДЗ 9-52 | физическая нагрузка | ЧСС | внутриклассовые корреляции | показатель наследуемости 0,58 |
| Л. Сергиенко, 1975 | 24 пары МЗ 26 пар ДЗ 12-17 | физическая нагрузка | ЧСС | внутриклассовые корреляции | показатель наследуемости 0,58 |
| П. Звольский и др. (P.Zvolskyetal.), 1976 | 15 пар МЗ 19 пар ДЗ 18-19 | покой, напряжение (стресс | частота пульса | внутриклассовые корреляции | наследуемость не зависит от ситуации; Н в диапазоне 0,6—0,9 |
| М. Фейнлейб и др. (М. Feinleib et al.), 1977 | 250 пар МЗ 264 пары ДЗ 42-56 | Покой | давление | дисперсионный анализ | показатели наследуемости для систолического давления 0,60; для диасто-лического — 0,61 |
| Е. Соколов с соавт., 1980 | 24 пары МЗ 15 пар ДЗ 17-53 | покой (1), напряжение (2), отдых (3) | давление, ЧСС | внутриклассовые корреляции | показатели наследуемости для систолического давления; 0,47, 0,81, 0,81; для диастолического — 0,73, 0,77, 0,53; для ЧСС: 0,20, 0,78, 0,70 |
| Б. Кочубей, 1983 | 22 пары МЗ 21 пара ДЗ 17-29 | звуки 80 дБ, 100 дБ, 1000 Гц | ЧСС, интенсивность привыкания | дисперсионный анализ | наследуемость ЧСС на звук 105 дБ (G составляет 45%) и интенсивности привыкания на звук 80 дБ (G — 42%) |
| Р. Сомсен и др. (R. Somsen et al.), 1985 | ПпарМЗ 11 пар ДЗ 15-18 | задачи на время реакции, вычисления в | ЧСС | внутриклассовые корреляции | при вычислении в уме МЗ близнецы более похожи, чем ДЗ |
| Б. Дито (В. Ditto), 1987 | 36 пар сиблингов | покой, вычисление в уме, действие | ЧСС | метод подбора моделей | большее сходство ЧСС и показателей давления в покое; меньшее сходство реактивных изменении |
| Б. Дито (В. Ditto), 1988 | 40 пар МЗ 40 пар ДЗ | задачи разного типа | ЧСС, давление | внутриклассовые корреляции | наследуемость реактивных изменений показателей от 0 до 0,80 в зависимости от характера задачи |
| Д. Бумсма и др. (D. Boomsma et al.), юоп Т990. Бушар (Т. Bouchard etal.), 1990 | 70 пар МЗ 90 пар ДЗ 1Л.17 34 пары МЗ, росших вместе 49 пар разлученных МЗ | покой, задачи покой | ЧСС, давление респираторно-синусная аритмиясистолическ | метод подбора моделей внутриклассовые корреляции | наследуемость в покое 25%; при выполнении задачи 50% все показатели наследуемы для давления rМЗ разлученных 0,6 для ЧСС rМЗ разлученных 0,4 |
| давление, ЧСС |
Общим
результатом
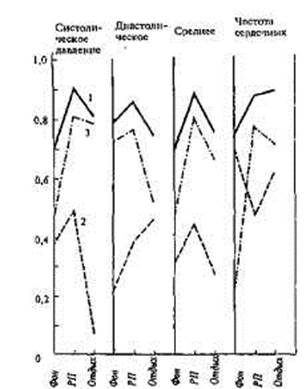
ДС.Ш1СНИС *flWlctmc сокращении ется тот факт, что все перечисленные показатели обнаружили наибольшую величину наследуемости (по Хольцингеру) в условиях интеллектуальной деятельности, т.е. при психоэмоциональной нагрузке (рис. 15.1; 15.2),
В то же время отмечаются существенные различия в изменениях показателей наследуемости систолического и диастолического давления в зависимости от состояния организма. Для систолического давления Н в покое составляет 0,47, при нагрузке — 0,81, после нагрузки — 0,79; для диастолического — соответственно 0,73; 0,77; 0,53. Поскольку уровень
давления крови при на-
Рис. 15.1. Изменение коэффициентов внут- грузке достоверно повы-риклассовой корреляции в группе монози- шался, то можно считать, готных (rМЗ), дизиготных (rДЗ) близнецов и что изменчивость всех коэффициента Хольцингера (Н) для систо- компонентов, определяю-лического, диастолического и среднего дав- щих величину систоличес-ления и частоты сердечных сокращений в кого давления крови при покое (фон), во время психоэмоциональной эмоциональном напряже-нагрузки (РП) и через 10 мин отдыха (от- детерминируется ге-
дых). РП— рабочий период [по 141]. н
нетическими факторами.
1 - rМЗ; 2 - rДЗ; 3 -Н. Наследуемость механизмов
диастолического давления не обнаруживает столь значительной связи с функциональным состоянием организма, оставаясь высокой и в покое, и при нагрузке.
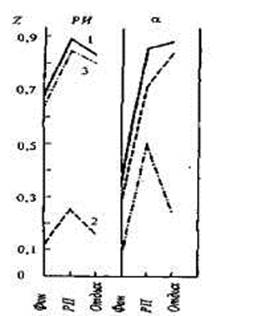
Наряду с этим в некоторых исследованиях не обнаружено столь отчетливых различий наследуемости давления крови, характерного для покоя и функциональной нагрузки. Примером служит исследование Д. Бумсма с соавторами, в котором изучалась природа межиндивидуальной вариативности уровня давления крови в зависимости от на-
пряженности ситуации. Давление регистрировали у 160 пар близнецов в возрасте 14-21 года в покое и при выполнении задач, включающих регистрацию времени реакции и вычисления в уме [200]. Результаты получились неоднозначными. Оценка наследуемости уровня систолического и диастолического давления у женщин была выше при нагрузках, у мужчин же увеличение наследуемости наблюдалось только для систолического давления. Более того, диастолическое давление мужчин при нагрузках зависело от генотипа в меньшей степени, чем в покое. В состоянии покоя в изменчивости систолического
давления у обоих полов и диас-
толического давления у женщин Рис. 15.2. Динамика коэффициентов
наблюдалось влияние системати- внутриклассовой корреляции в груп-
ческой среды, однако при функ- пе монозиготных (rмз), дизиготных
циональных нагрузках это влия- (rдз) близнецов и коэффициента
ние снижалось. Многомерный Хольцингера (Н) для реографичес-
дисперсионный анализ данных кого индекса (РИ) и времени кро-
позволил заключить, что и в по- венаполнения сосудов головного
кое, и при функциональных на- мозга (а) в покое(фон), во время
психоэмоциональной нагрузки (РП)
грузках природа генетических и 1А и д , г
VJ v v „ и через 10 мин отдыха (отдых) [по
средовых влияний остается неиз- \\\л
менной.
В целом ряде работ было об- 1- rМЗ; 2 - rДЗ; 3 - Н. наружено большее сходство реактивных изменений частоты сердечных сокращений в ответ на сенсорные стимулы и при нагрузках в парах МЗ близнецов по сравнению с ДЗ (табл. 15.3). В связи с этим широкое распространение получило мнение, что межиндивидуальная вариативность показателей частоты сердечных сокращений (ЧСС) при максимальной нагрузке зависит от наследственных факторов, тогда как индивидуальные особенности ЧСС в покое зависят, как правило, от факторов среды. Наряду с этим, однако, имеются и противоположные наблюдения.
Прежде, чем обсудить эти расхождения, следует указать, что сама по себе динамика ЧСС при выполнении задании разного рода имеет неоднозначный характер. При предъявлении задания и в ходе его выполнения возможно как ускорение, так и замедление ЧСС. В качестве
иллюстрации можно привести исследование Р.Сомсена со соавторами, в котором изучалось влияние генетических факторов на форму реактивных изменений ЧСС при выполнении разных заданий у 11 пар МЗ и 11 пар ДЗ близнецов [409]. Предварительно было установлено, что текущие изменения ЧСС имеют характерный паттерн, который зависит от особенностей задачи и этапа ее решения. Задания были связаны с регистрацией времени простой двигательной реакции и вычислением в уме. В заданиях на время реакции при предъявлении предупреждающего сигнала наблюдалось замедление ЧСС, затем ускорение, а перед предъявлением пускового сигнала — опять замедление. При этом авторы отмечают высокую межиндивидуальную вариативность паттерна изменений ЧСС, хотя у каждого испытуемого он оставался стабильным. Тем не менее достоверных различий между МЗ и ДЗ близнецами по данному признаку в этих условиях выявлено не было. В другом случае простая двигательная реакция перемежалась заданиями на вычисление в уме. С точки зрения авторов, в таком случае возникала более напряженная, стрессогенная ситуация. В этой ситуации сходство паттерна ЧСС-реакции оказалось достоверно больше у МЗ близнецов, чем у ДЗ. Предположительно, различия в наследуемости паттерна реактивности ЧСС могут иметь две причины. Во-первых, при больших нагрузках и стрессе показатели ЧСС оказываются более стабильными, поэтому низкая оценка наследуемости ЧСС-реакции при выполнении относительно простого задания может быть обусловлена большей ошибкой измерения. Вторая причина — различная наследуемость показателей, обусловленных активностью симпатической и парасимпатической систем.
Изменения показателей работы сердечно-сосудистой системы находятся под контролем симпатического и парасимпатического отделов вегетативной нервной системы, причем те и другие влияния по-разному действуют на физиологические показатели, в частности на ЧСС, Парасимпатическая активация вызывает замедление сердечного ритма, которое сопровождает, например, ориентировочную реакцию, возникающую при предъявлении новых умеренных по интенсивности стимулов, или просто внимательное их рассматривание. Известно, что ЧСС в задаче на время реакции также определяется в основном активностью парасимпатической системы, а при задаче, связанной с вычислениями в уме, — в большей степени симпатической системы. Симпатическая активация, напротив, ведет к увеличению частоты сердечных сокращений и повышению мышечного напряжения. Данные аффекты тесно связаны, поэтому любое мышечное напряжение сопровождается усилением ЧСС. Значимость этой связи наиболее выпукло обозначена в теории кардиосома-тического сопряжения Р. Обриста, в соответствии с которой ритм сердца и уровень мышечного напряжения взаимообусловлены: при ослаблении мышечного напряжения замедляется ритм сердца, и наоборот.
Полученные Р. Сомсеном данные позволяют предположить, что эффекты симпатической активации в большей степени обусловлены генотипом, чем аналогичные эффекты парасимпатической системы.
К сходным выводам пришел и Б.И. Кочубей [84] в процессе изучения изменений ЧСС в ситуациях ориентировочно-исследовательской и ориентировочно-оборонительной реакций на звуки разной интенсивности у 22 пар МЗ и 21 пары ДЗ близнецов.
Как уже отмечалось, для ориентировочной реакции характерно замедление сердечного ритма. Оно обусловлено действием блуждающего нерва (парасимпатическая система), а психологическим выражением соматических эффектов является «обращенность» человека вовне. Для оборонительной реакции, наоборот, характерно учащение ритма, обусловленное симпатической активацией. Оно свидетельствует об установке испытуемого на избегание стимула. По данным Б.И. Кочубея, стимул 80 дБ вызывал уменьшение ЧСС, а при привыкании ориентировочной реакции наблюдалось относительное учащение ритма. Звук 105 дБ, напротив, сопровождался увеличением ЧСС, а при его повторении отмечалось относительное снижение ЧСС. Значительный вклад генетических факторов наблюдался при увеличении ЧСС в ответ на тон 105 дБ (оборонительная реакция — ОбР) и отсутствовал в изменениях ЧСС в ответ на тон 80 дБ (ориентировочная реакция — Ор). Величина привыкания, наоборот, была обусловлена генотипом только при тоне 80 дБ. Таким образом, генетические влияния наблюдались в ситуациях, когда ведущую роль играла симпатическая регуляция сердечного ритма (угашение Ор и первая реакция при ОбР), тогда как в ситуациях, характеризующихся преобладанием парасимпатических влияний, вариативность ЧСС определялась преимущественно средовыми влияниями.
При изучении деятельности автономной нервной системы используются показатели, отражающие взаимодействие и меру согласованности в работе сердечно-сосудистой и дыхательной систем организма. Одним из таких показателей является респираторная синусная аритмия (РСА). Она отражает циклические изменения ЧСС, сопровождающие дыхание. ЧСС обычно увеличивается при выдохе и уменьшается при вдохе. Чем больше амплитуда изменения ЧСС, тем выше РСА. Считается, что высокий уровень РСА свидетельствует о хорошем контроле ЧСС со стороны парасимпатической нервной системы (вагусный контроль). Выраженный вагусный контроль ЧСС и, следовательно, высокая РСА рассматриваются как признак хорошей регуляции в деятельности сердечно-сосудистой и автономной нервной систем.
Д. Бумсма с коллегами исследовали влияние средовых и генотипи-ческих факторов на межиндивидуальную вариативность РСА в покое и при выполнении задач двух типов — на время реакции и вычисления в уме [201]. Методом подбора моделей было установлено, что при выполнении задач приблизительно 50% общей дисперсии объяснялось генетическими факторами, тогда как в покое — около 25%. Описанию данных более всего соответствовала генотип-средовая модель,
включающая случайный средовой и аддитивный генетический компоненты дисперсии. Влияния систематической среды на дисперсию РСА обнаружено не было. Было также установлено, что дисперсия РСА в покое и при выполнении задач имеет общую генетическую основу, т.е. определяется действием одних генетических факторов.
В работе эстонских исследователей [132, гл. VI] на большой выборке близнецов (153 пары) была установлена наследственная обусловленность деятельности систем кровообращения и дыхания лишь в условиях больших физических нагрузок. В состоянии покоя и при умеренных усилиях средовые влияния преобладали над генотипическими.
Было также установлено значительное влияние факторов генотипа на межиндивидуальную вариативность некоторых параметров кровообращения и максимального потребления кислорода (МПК) при выполнении спортивных движений. Исследования МПК оказываются наиболее интересными. Этот показатель (от которого в решающей мере зависит успешность в циклических видах спорта) одинаков в разных этнических группах, не меняется в онтогенезе, мало тренируется и оказывается высоко генетически детерминированным [310]. Если учесть, что спортсмены международного класса имеют показатели МПК, значительно превышающие их среднепопуляционную величину, то, по-видимому, индивидуальный уровень МПК может служить информативным признаком при прогнозировании спортивной успешности.
Дата добавления: 2015-07-18; просмотров: 1587;