ПОТЕНЦИАЛЫ МОЗГА, СВЯЗАННЫЕ С ДВИЖЕНИЕМ
Среди событийно-связанных потенциалов особое место занимают потенциалы мозга, связанные с движением (ПМСД). Изучение ПМСД позволяет выявить скрытую последовательность процессов, происходящих в коре мозга при подготовке и выполнении движения, и хронометрировать эти процессы, т.е. установить временные границы их протекания [104, 237].
Впервые комплекс колебаний, отражающий процессы подготовки, выполнения и оценки движения, был зарегистрирован в 60-х годах. Оказалось, что движению предшествует медленное отрицательное колебание — потенциал готовности (ПГ). Он начинает развиваться за 1,5-0,5 мс до начала движения. Этот компонент регистрируется преимущественно в центральных и лобно-центральных отведениях обоих полушарий. Принято считать, что ПГ возникает в моторной коре и связан с процессами планирования и подготовки движения. Он относится к классу медленных негативных колебаний потенциала мозга, возникновение которых объясняют активацией нейрональных элементов соответствующих участков коры.
За 500-300 мс до начала движения ПГ становится асимметричным — его максимальная амплитуда наблюдается в прецентральной области, контралатеральной движению. Примерно у половины взрослых испытуемых на фоне этого медленного отрицательного колебания незадолго до начала движения регистрируется небольшой по амплитуде положительный компонент — Р1. Следующее по порядку быстро нарастающее по амплитуде отрицательное колебание N2, так называемый моторный потенциал (МП), начинает развиваться за 150 мс до начала движения и достигает максимальной амплитуды над областью, в которой находятся корковые центры управления движу-
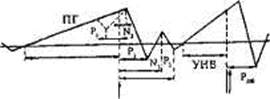
Рис. 14.6. Схема потенциалов, связанных с реализацией двигательного действия.
Одной вертикальной стрелкой обозначено начало движения, двумя — сигнал обратной связи. Компонент Рь обозначен пунктирной линией, поскольку выделяется не у всех испытуемых. Одна стрелка — начало движения, две — сигнал обратной связи [104]. ПГ — потенциал готовности; УНВ — условная негативная волна.
щейся конечностью. Примерно через 200 мс после начала движения возникает положительный компонент Р2. Завершается этот комплекс потенциалов появлением компонентов N3 иР3 (рис. 14.6).
Еще один электрофизиологический феномен по своей сути близок потенциалу готовности. Речь идет об отрицательном колебании потенциала, регистрируемого в передних отделах коры мозга в период между действием предупреждающего и пускового (требующего реакции) сигналов. Это колебание имеет ряд названий: Е-волна, волна ожидания, условная негативная волна (УНВ). Е-волна возникает после предупредительного сигнала, ее длительность растет с увеличением интервала между первым и вторым стимулами. Амплитуда Е-волны возрастает прямо пропорционально скорости двигательной реакции на пусковой стимул. Она увеличивается при напряжении внимания и повышении волевого усилия, что свидетельствует о связи этого электрофизиологического явления с механизмами произвольной регуляции двигательной активности и поведения в целом.
Обнаруживая индивидуальную специфичность и стабильность, эти потенциалы дают возможность оценить вклад генетических факторов в изменчивость биоэлектрических коррелятов индивидуальных особенностей организации и построения движений.
Генетическое исследование ПМСД было проведено С.Б. Малыхом [104] на близнецах 18-30 лет (по 25 пар МЗ и ДЗ близнецов). Особенность этой работы — изучение генетической обусловленности ПМСД в трех психологически различающихся ситуациях: в первой требовалось произвольно (без предварительного сигнала) нажимать на кнопку; во второй и третьей надо было прогнозировать появление звуковых стимулов, организованных в ряд с фиксированной последовательностью и разной вероятностью. Испытуемому сообщалось, что звуки в правом и левом наушниках организованы в блоки, а именно:
звук слева может встречаться по 2 или 3 раза подряд, а справа — по 2 или 4 раза подряд. Надо было предсказать появление звука в правом или левом наушнике, сообщая о характере прогноза нажатием на одну из двух кнопок, расположенных справа. Описанная организация стимульного ряда позволяла получать прогноз событий, наступающих с вероятностьюр = 1,0 ир = 0,5.
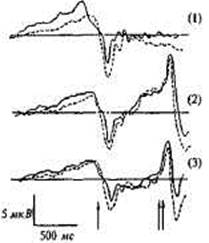
ПМСД получали усреднением ЭЭГ, зарегистрированной монопо-лярно в зонах F3, F4, C3, С4. Для каждого из четырех отведений у одного испытуемого было получено по три потенциала: при простых произвольных движениях без внешнего сигнала (1), при нажатии кнопки в
ситуации прогноза двух равноверо- Рис. 14.7. ПМСД, УНВ и Р300, ятных сигналов (2) и в ситуации зарегистрированные в отведении заведомо истинного прогноза (3). С3 у пары МЗ близнецов в трех эк-
В ситуации (1) анализировались спериментальных ситуациях [104]. потенциал готовности (ПГ), ком- Одной стрелкой отмечено начало дви-поненты Рь N2, Р2, N3, Р3 отра- жения, двумя — сигнал обратной жавшие соответственно процессы связи. подготовки, реализации и оценки
произведенного движения, В ситуациях (2) и (3) помимо указанных компонентов ПМСД регистрировалась условная негативная волна (УНВ), появлявшаяся между нажатием кнопки и ожидаемым сигналом обратной связи (т.е. звуком, который подтверждал или не подтверждал прогноз), сменявшаяся после предъявления звука положительным компонентом Р300 (рис. 14.7). Два последних компонента отражают процессы ожидания и оценки прогноза. Генетический анализ включал оценку внутрипарного сходства амплитуд и латентных периодов всех выделенных компонентов с помощью внутриклассовой корреляции.
Анализ влияний генотипа и среды на изменчивость потенциалов мозга, связанных с реализацией двигательного действия, дал достаточно пеструю картину (табл. 14,4). Для того чтобы выделить наиболее общие тенденции, по каждому признаку был подсчитан процент тех позиций, в которых обнаруживался генетический контроль. Оказалось, что в амплитудных параметрах генетический контроль обнаруживается чаще (63,2% всех случаев), чем во временных (21%). Правда, латентности компонентов ПМСД измеряются менее надежно,
чем в сенсорных ВП, из-за отсутствия внешнего сигнала, от которого обычно ведется счет времени. Возможно, это и сказалось на оценке наследуемости.
Выявились также определенные межзональные различия: менее всего генетический контроль выступает в показателях ПМСД левой лобной области (26,3%), в остальных трех зонах он выше (от 44,7 до 50%).
Наиболее важным представляется тот факт, что вклад генетической и средовой составляющих существенно меняется в зависимости от характера деятельности испытуемого. Чаще всего влияние генотипа обнаруживается в ситуации прогноза равновероятных событий (53,6% всех исследованных признаков), реже всего (20%) — при простых произвольных движениях, и промежуточное положение (46,4%) занимает ситуация с заведомо истинным прогнозом. Таким образом, при осуществлении прогнозирования генетический контроль обнаруживается чаще, чем при простых произвольных движениях, не являющихся ответом на внешний стимул и осуществляемых по субъективной команде (см. табл. 14.4).
Закономерно возникает вопрос: почему ПМСД, сопровождающие по фенотипическому проявлению одно и то же движение, обнаруживают столь разную степень генетической обусловленности? Объяснение этого факта видится в разном функциональном значении данного движения в изучаемых ситуациях. При простом произвольном нажатии кнопки испытуемый сам инициирует движение. Иначе говоря, в соответствии с инструкцией цель действия — само нажатие на кнопку. Во второй ситуации целью является прогноз, а движение становится лишь средством достижения этой цели. Поскольку двигательные акты в первом и втором случаях различаются, прежде всего по степени произвольности и осознаваемости и, следовательно, по уровню нейрофизиологического обеспечения, постольку изменение роли генотипа в вариативности соответствующих ПМСД можно объяснить, по-видимому, отражением в их параметрах активности разных функциональных систем.
Конфигурация ВП, а также их основные количественные параметры относятся к числу индивидуально устойчивых особенностей человека, что дает основания изучать роль генотипа и среды в происхождении межиндивидуальной вариативности по этим признакам.
В целом имеющиеся в литературе данные свидетельствуют о существенном влиянии генотипа на межиндивидуальную вариативность вызванных и событийно-связанных потенциалов. Генетическую обусловленность обнаруживают сенсорные (зрительные и слуховые) ВП, а также потенциалы мозга, связанные с движением.
Таблица 14.4
Наличие генетической составляющей в вариативности амплитудно-временных характеристик компонентов
потенциала в трех экспериментальных ситуациях [104]
| Амплитуды | Временные | i характеристики | |||||||||||||||||||||||||
| Комп | |||||||||||||||||||||||||||
| о- | |||||||||||||||||||||||||||
| нент | F3 | С3 | C4 | F3 | С3 | С4 | F3 | С3 | C4 | F3 | C3 | С4 | F3 | C3 | С4 | F3 | С3 | C4 | |||||||||
| ы | |||||||||||||||||||||||||||
| ПГ | + | + | + | + | + | + | + | + | + | ||||||||||||||||||
| N2 | + | + | + | + | + | + | + | + | |||||||||||||||||||
| P2 | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | |||||||||||
| N3 | + | + | + | + | + | + | + | ||||||||||||||||||||
| Рз | + | + | + | + | + | + | + | + | |||||||||||||||||||
| УНВ | — | — | — | — | + | + | + | + | + | — | — | — | — | + | + | + | |||||||||||
| P300 | — | — | — | — | + | + | + | + | + | + | + | — | — | — | — | + | |||||||||||
Примечание. Прочерки означают отсутствие компонента, которых влияют генотипические факторы.
знаком
выделены признаки, на межиндивидуальную вариативность
Однако соотношение генетических и средовых детерминант в изменчивости ВП может меняться в зависимости от модальности стимула, его интенсивности и семантических особенностей. Более отчетливо она проявляется при возрастании интенсивности стимула. В ответах на семантические стимулы влияние генотипа сказывается меньше, чем в ответах на элементарные сенсорные стимулы. Привлечение непроизвольного (ориентировочная реакция) и произвольного внимания к стимулу также способствует увеличению доли генетической составляющей вариативности ранних компонентов ВП. Наряду с этим в параметрах поздних компонентов ВП при мобилизации внимания обнаруживается влияние систематической среды.
Степень генетической обусловленности ПМСД зависит от характера деятельности испытуемого. Она меньше, когда движение является целью, и больше, когда оно выступает в качестве средства для достижения цели.
Все это означает, в свою очередь, что, во-первых, принадлежность исследуемого признака к так называемому биологическому (в данном случае нейрофизиологическому) уровню не означает его безусловной генетической детерминированности и, во-вторых, психологические факторы, включаясь в реализацию такого признака, могут, существенно не меняя его фенотипического проявления, изменять детерминанты его фенотипической дисперсии,
Глава X V
ГЕНОТИП-СРЕДОВЫЕ СООТНОШЕНИЯ В ИЗМЕНЧИВОСТИ ПОКАЗАТЕЛЕЙ ВЕГЕТАТИВНЫХ РЕАКЦИЙ
Традиционным объектом психофизиологических исследований являются показатели функционирования физиологических систем организма (сердечно-сосудистой, дыхательной, мышечной, выделительной), которые закономерно изменяются при психической деятельности. Как правило, показатели активности этих систем отличаются индивидуальной специфичностью и достаточно устойчивой воспроизводимостью при повторных регистрациях в одинаковых условиях, что дает основание ставить вопрос о роли генотипа в происхождении этих различий.
Исследования генетических основ изменчивости вегетативных функций несистематичны, проведены в разной логике и с различными методами регистрации тех или иных реакций, а потому объединить их в единую систему знаний о происхождении индивидуальных различий этого уровня в структуре индивидуальности весьма сложно. Такое положение дел, естественно, не может не вызвать сожаления,
Однако соотношение генетических и средовых детерминант в изменчивости ВП может меняться в зависимости от модальности стимула, его интенсивности и семантических особенностей. Более отчетливо она проявляется при возрастании интенсивности стимула. В ответах на семантические стимулы влияние генотипа сказывается меньше, чем в ответах на элементарные сенсорные стимулы. Привлечение непроизвольного (ориентировочная реакция) и произвольного внимания к стимулу также способствует увеличению доли генетической составляющей вариативности ранних компонентов ВП. Наряду с этим в параметрах поздних компонентов ВП при мобилизации внимания обнаруживается влияние систематической среды.
Степень генетической обусловленности ПМСД зависит от характера деятельности испытуемого. Она меньше, когда движение является целью, и больше, когда оно выступает в качестве средства для достижения цели.
Все это означает, в свою очередь, что, во-первых, принадлежность исследуемого признака к так называемому биологическому (в данном случае нейрофизиологическому) уровню не означает его безусловной генетической детерминированности и, во-вторых, психологические факторы, включаясь в реализацию такого признака, могут, существенно не меняя его фенотипического проявления, изменять детерминанты его фенотипической дисперсии,
Глава X V
ГЕНОТИП-СРЕДОВЫЕ СООТНОШЕНИЯ В ИЗМЕНЧИВОСТИ ПОКАЗАТЕЛЕЙ ВЕГЕТАТИВНЫХ РЕАКЦИЙ
Традиционным объектом психофизиологических исследований являются показатели функционирования физиологических систем организма (сердечно-сосудистой, дыхательной, мышечной, выделительной), которые закономерно изменяются при психической деятельности. Как правило, показатели активности этих систем отличаются индивидуальной специфичностью и достаточно устойчивой воспроизводимостью при повторных регистрациях в одинаковых условиях, что дает основание ставить вопрос о роли генотипа в происхождении этих различий.
Исследования генетических основ изменчивости вегетативных функций несистематичны, проведены в разной логике и с различными методами регистрации тех или иных реакций, а потому объединить их в единую систему знаний о происхождении индивидуальных различий этого уровня в структуре индивидуальности весьма сложно. Такое положение дел, естественно, не может не вызвать сожаления,
так как особенности функционирования вегетативной системы тесно связаны с динамикой функциональных состояний человека, ее показатели используются при изучении эмоционально-волевой сферы и интеллектуальной деятельности человека [436, 84, 203].
Дата добавления: 2015-07-18; просмотров: 2059;