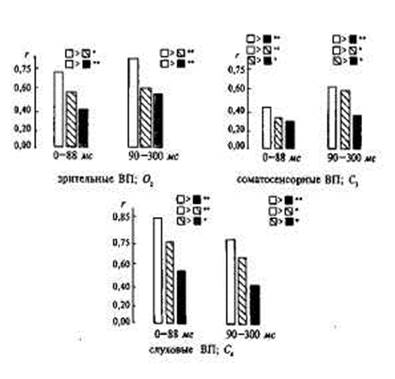
Вкаждой изтрех групп (МЗ, ДЗ, HP)волновую форму попарно коррелировали, а затемусредняли (М6|. П - МЗ; Q- ДЗ; ■ - HP; • - р > 0,05; •• - р > 0,01.
стью, в 2 раза превышающей субъективный порог. По волновой форме соматосенсорных ВП статистически достоверных различий между МЗ и ДЗ получено не было. Изучение амплитудных показателей позволило выявить генетические влияния в изменчивости соматосенсорных ВП, но они все равно оказались меньше, чем в зрительных и слуховых ВП.
Следующей характеристикой стимула является его интенсивность. В каждой модальности стимулы могут быть слабыми, умеренными и сильными, соответственно амплитуда ВП изменяется в зависимости от интенсивности стимула. Если стимулы (вспышка, звуковой тон) слабые, то ВП имеют небольшую амплитуду. При усилении стимула амплитуда компонентов ВП возрастает, но только до определенного предела, различного у разных индивидов. По данному признаку они делятся на «уменьшителен» и «увеличителей»: у первых этот предел наступает раньше, чем у вторых.
Закономерно возникает вопрос: зависят ли проявления генетического контроля ВП от силы раздражителя? Судя по некоторым данным, генетическая обусловленность амплитуды проявляется по-раз-
ному в ответах на стимулы различной силы [ 132, гл. IV]. В слуховых ВП увеличение интенсивности раздражителя приводит к более отчетливому проявлению генотипического контроля амплитуд. Различия в факторах, формирующих изменчивость ВП на тоны 80 дБ и 105 дБ, анализировались Б.И. Кочубеем [84]. Влияние генотипа отчетливо сказывалось в амплитудах компонентов N1 Р2 (интервал 100-200 мс): показатели наследуемости составили 0,46 и 0,79 в реакции на тон 80 дБ и 0,58 и 0,81 в реакции на тон 105 дБ.
Эта проблема исследовалась также применительно к зрительным и слуховым ВП в связи с изучением генетического контроля феномена «увеличения—уменьшения» [217, 218]. Изучение внутрипарного сходства динамики ВП при изменении интенсивности стимулов (вспышки) у 33 пар МЗ и 34 пар ДЗ близнецов дало внутриклассовые корреляции, свидетельствующие об увеличении наследуемости амплитуды ВП на световые стимулы по мере возрастания яркости стимула. В табл. 14.1 приведены данные для амплитудных показателей, вычисленных в интервале 76-112 мс. Из этих данных следует, что по
Таблица 14.1
Наследуемость амплитудных параметров зрительных ВП
| Амплитудные | Коэффициенты | Коэффициенты | Показатель |
| показатели | корреляции МЗ | корреляции ДЗ | наследуемости |
| При разных интен- | |||
| сивностях стимула: | |||
| 1-й уровень | -0,20 | -0,21 | 0,00 |
| 2-й уровень | 0,31 | 0,07 | 0,25 |
| 3-й уровень | 0,40 | 0,02 | 0,39 |
| 4-й уровень | 0,51 | -0,04 | 0,52 |
| Средние амплитуды: | |||
| Р1 00 -N140 | 0,59 | 0,36 | 0,35 |
| N140 - P200 | 0,57 | 0,10 | 0,52 |
| Крутизна наклона кри- | |||
| вой при росте интен- | |||
| сивности стимула: | |||
| P1 00 -N140 | 0,51 | -0,04 | 0,52 |
| N140 - P200 | 0,56 | -0,10 | 0,60 |
Примечание. Таблица составлена по данным М. Бухсбаума [217]. Уровни интенсивности стимула даны по возрастающей; наследуемость оценивалась по Хольцингеру (именно этот коэффициент использовал М. Бухсбаум).
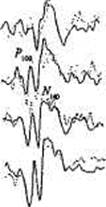
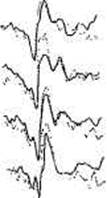
Рис. 14.3. Зрительные ВП двух пар МЗ близнецов в ответ на четыре уровня световой интенсивности.
В каждой паре ВП одного близнеца представлен сплошной линией, второго — пунктирной. Отметьте сходство латентностей пиков, волновой формы и изменений по мере роста интенсивности стимула. В паре 1 — слева — компонент Р100 N140 заметно возрастает по мере роста интенсивности стимула (augmenting), в паре 2 тот же компонент уменьшается (reducing) [217].
мере возрастания интенсивности влияние генотипа в амплитуде зрительных ВП проявляется все более отчетливо.
Как подчеркивает автор, на втором или третьем уровне интенсивности происходило деление испытуемых на «увеличителей» и «умень-шителей», т.е. сохраняющих или изменяющих тип реагирования на усиление интенсивности стимула (рис. 14.3). Внутрипарное сопоставление наклона прямой, отражающей зависимость амплитуды ВП от силы раздражителя, выявило существенно большее сходство МЗ близнецов по сравнению с ДЗ. Таким образом, установлено, что крутизна наклона прямой, отражающей зависимость амплитуды ВП от силы раздражителя, в значительной степени контролируется генотипом.
Генетическая детерминация индивидуальных различий по феномену «увеличения—уменьшения» подтвердилась и в семейных исследованиях. Было установлено, что межиндивидуальные различия по данному признаку на 51-68% объясняются генетическими влияниями. Кроме того, по этому признаку наблюдается ассортативность в подборе супружеских пар, корреляции между супругами составляют 0,32 [218].
Психофизиологическая сущность феномена «увеличения—уменьшения» не совсем ясна и требует более глубокого изучения. Не исключено, что в его
основе лежат некоторые общебиологические механизмы, отражающие «пропускную способность» нервной системы при обработке информации. По-видимому, такие механизмы ограничивают способность человека и животных адекватно реагировать на стимулы при слишком значительном увеличении их интенсивности. Ограничения в «пропускной способности» сенсорных каналов ЦНС, накладываемые указанными механизмами, индивидуально вариативны, и в их межиндивидуальную вариативность вносят существенный вклад генетические факторы.
Так или иначе, но в изменчивости и зрительных, и слуховых ВП усиление интенсивности стимула приводит к более отчетливому проявлению генетической обусловленности амплитудных параметров.
Кроме модальности и интенсивности, стимулы могут различаться и другими особенностями — качественными и количественными. Например, зрительные стимулы могут иметь одинаковые физические параметры (освещенность), но разный рисунок и/или содержание. В частности, зрительные стимулы в виде шахматных полей, имея одинаковую общую освещенность, будут иметь разный вид, определяемый размером ячейки. При этом ВП на шахматные паттерны с ячейками разных размеров также будут существенно различаться [134].
Изучение генетической обусловленности параметров ВП на сменяющие друг друга шахматные поля с разными ячейками (обращаемый шахматный паттерн) было проведено К.Б. Булаевой с соавторами [219] в семейном исследовании. При анализе сходства между родителями и детьми, а также между сиблингами была установлена наследуемость амплитуд и латентностей ранних компонентов (интервал 0-100 мс) зрительных ВП на обращаемый шахматный паттерн. Показатели наследуемости для разных компонентов ВП варьирует от 0,28 до 0,88.
Вызванные потенциалы изменяют свои параметры при предъявлении стимулов не только разной формы, но и разного содержания. Влияние содержательных особенностей стимула на генотип-средовые соотношения в изменчивости ВП исследовали Т.М. Марютина и Т.Г. Ивошина [109] у взрослых близнецов применительно к 7 вариантам стимулов (рис. 14.4).
По порядку предъявления это были: вспышка; симметричная геометрическая фигура, не имеющая названия; комбинация букв ДМО; хаотический набор элементов, из которых складывалось изображение дома; слово ДОМ; рисунок дома; шахматное поле с ячейкой 20 минут. ВП регистрировались монополярно из 6 зон (02, C2, T5, Т6, F3, F4 по системе «10-20»). В зависимости от особенностей стимула менялись амплитуды и латентности компонентов ВП. Оценка внутрипарного сходства проводилась по волновой форме, латен-тностям и амплитудам всех компонентов ВП.
По совокупности полученных данных было подсчитано 336 коэффициентов наследуемости, для сравнения ВП на разные стимулы ис-

пользовалась обобщенная характеристика — процент тех позиций, по которым внутри-парное сходство МЗ и ДЗ близнецов различалось статистически достоверно. Такой прием позволил выявить интересный факт: максимум генетических влияний был установлен для параметров ВП на вспышку и шахматное поле (60 и 62% соответственно), минимум — для ответов на семантические стимулы (рисунок дома и слово ДОМ — 31 и 29%). Стимулы 2, 3, 4 получили по 48, 50 и 45% соответственно.
Разложение фенотипичес-кой дисперсии латентных периодов ВП обнаружило значительную долю генетической изменчивости в ответах на вспышку и шахматное поле. В ответах на семантические стимулы, напротив, отчетливо выступает влияние систематической среды (табл. 14.2). Таким образом, роль генетической и средовой изменчивости в формировании индивидуальных особенностей зрительных ВП существенно зависит от содержания стимула. Предполагается, что в основе этих различий лежит разное физиологическое обеспечение элементарных сенсорных процессов и перцептивной деятельности, формирующейся в ходе освоения социального опыта.
Итак, степень генетической обусловленности ВП проявляется по-разному в зависимости от модальности стимула, его интенсивности, графических и семантических особенностей. Иначе говоря, даже в психофизиологическом феномене — ВП, принадлежащем, условно говоря, к «индивидному» биологическому уровню в структуре индивидуальности, соотношение генетических и средовых влияний зависит от особенностей переработки информации, т.е. от когнитивного уровня.
Рис. 14.4. Зрительные ВП затылочной области в парах МЗ и ДЗ близнецов. Толстой линией обозначены ВП одного близнеца, тонкой — ВП другого близнеца. Числа — коэффициенты корреляции, иллюстрирующие динамику внутрипарного сходства ВП по волновой форме в целом (0—512 мс) в зависимости от вида стимула [132, гл. IV].
Таблица 14.2
Коэффициенты внутриклассовой корреляции и разложение (в %)
фенотипической дисперсии латентных периодов компонентов ВП
затылочной области при предъявлении разных стимулов [132, гл. IV]
| Стимулы | Компоненты | rM3 | Гдз | VА | VD | VC | VW | х2(n= 2) | P | |
| -Г1 ПО N120 P1 40 N200 P250 | 0,73* 0,56* 0,68* 0,80* 0,67 | 0,45 -0,32 0,07 -0,03 0,26 | 67 63 | 23 69 79 | — | 33 77 31 21 | 2,73 0,84 0,40 1,75 1,35 | 0,25 0,50 0,75 0,50 0,50 | 0,50 0,75 0,90 0,25 0,75 | |
| P1 00 N120 Р1 40 N200 P250 | 0,31 0,67* 0,65* 0,88* 0,18 | -0,07 0,18 0,19 0,48 0,46* | 28 85 | — | 47 15 100 | 3,44 6,46 0,44 1,35 | 0,10 0,025 0,75 0,50 | 0,25 0,90 0,75 | ||
| P1 00 N120 P1 40 N200 P250 | 0,55* 0,35 -0,06 0,49* 0,37 | -0,24 -0,38 -0,14 0,48 0,08 | — | 23 51 | 67 77 49 50 | 4,43 0,12 0,73 0,69 0,08 | 0,10 0,90 0,50 0,50 0,95 | 0,25 0,95 0,75 0,75 0,99 | ||
| P1 00 N120 P1 40 N200 P250 | 0,32 0,52 0,27 0,31 0,45 | 0,03 0,31 0,21 0,65** 0,56* | — | 26 30 55 44 | 74 74 70 45 56 | 2,80 0,17 3,23 2 65 1,72 | 0,10 0,90 0,10 0 95 0,25 | 0,25 0,95 0,25 0,50 0,50 |
Примечание. * —р < 0,05; ** —р < 0,01; полужирным шрифтом выделены те rМЗ, которые статистически значимо отличаются от rДЗ.
ВЛИЯНИЕ ОСОБЕННОСТЕЙ ЗАДАЧИ
Известно, что субъективное отношение испытуемого к стимулу, например сосредоточение внимания и отвлечение внимания, меняет рисунок и параметры ВП. Возникает вопрос: как влияет этот субъективный фактор на природу межиндивидуальной вариативности ВП? Коль скоро важную роль в формировании механизмов произвольной регуляции играет жизненный опыт индивида, то есть основания предполагать определенное влияние средовых факторов на параметры ВП, полученные в подобных условиях.
В первых исследованиях внимания методом ВП использовались простые поведенческие модели, например счет стимулов. При этом было установлено, что произвольное привлечение внимания испытуемых к стимулу сопровождается увеличением амплитуды компонентов ВП и сокращением их латентностей. Аналогичные изменения параметров ВП характерны и для ориентировочной реакции (непроизвольного привлечения внимания к стимулу). Напротив, по мере привыкания испытуемого к стимулу наблюдается снижение амплитуд и увеличение латентностей ВП.
Исследование ВП у близнецов 8—12 лет в ситуации ориентировочной реакции и при счете вспышек, предъявляемых с частотой 1 раз в 5 с, показало, что при привлечении внимания испытуемых к стимулу по сравнению с ситуацией привыкания к стимулу имеет место значительное увеличение генотипического контроля в вариативности амплитуд и латентностей большинства компонентов ВП [132, гл. IV]. В ситуации привыкания только 17% показателей обнаружили зависимость от генотипа, при счете вспышек генетический контроль обнаруживался в 43% случаев, в условиях ориентировочной реакции — в 69%. Генетико-дисперси-онный анализ показал, что при привлечении внимания испытуемых к стимулу наряду с генетическими влияниями присутствуют влияния систематической среды преимущественно в латентностях поздних компонентов (в интервале 200—300 мс). При пассивном отвлечении внимания от стимула в показателях ВП увеличивается доля влияний случайной среды.
Сходные результаты были получены при изучении слуховых ВП на тоны 1000 Гц интенсивностью 80 дБ в условиях ориентировочной реакции [80]. В амплитудах ранних компонентов ВП N1, Р2 (в интервале 100-200 мс) отчетливо выступает влияние генотипа, в амплитудах более поздних компонентов ВП N2, Р3 (в интервале 200—350 мс) — влияние факторов систематической среды.
Предполагается, что изменение параметров ВП при привлечении внимания к стимулу в условиях ритмической стимуляции происходит за счет общего усиления неспецифической подкорковой активации. Это позволяет предположить, что более жесткий генотипический контроль параметров ВП в ситуации внимания связан с модифицирующим действием восходящей подкорковой активации.
В онтогенезе формирование механизмов произвольной регуляции перцептивной деятельности, в том числе внимания, происходит постепенно и определяется совершенствованием механизмов управляемой корковой активации, механизмы которой в общих чертах складываются только к 9-10 годам [143]. При таком длительном периоде созревания логично ожидать определенного вклада средовых воздействий в изменчивость механизмов, обеспечивающих процессы управляемой корковой активации. Последнее отчасти объясняет, почему ВП в ситуациях привлечения внимания к стимулам, отражая особенности активации в этих условиях, обнаруживают, с одной стороны, возросшую степень генетической обусловленности, а с другой — отчетливую зависимость от влияний систематической среды.
21-1432 321
Таблица 14.3
Наследуемость параметровслуховых ВП вситуации равновероятного представлениястимулов (oddball paradigm) [no: 431]
| Автор, | Испытуемые, | Параметры | Использо- | Основные |
| год публи- | возраст | ВП | ванные | результаты |
| кации | (в годах) | методы | ||
| оценивания | ||||
| В. Севилло | 6 пар МЗ | латентные | критерий | для Nb Р2: конкор- |
| (W.Survillo), | 6 пар HP | периоды | Манна- | дантность МЗ и HP |
| 9-13 | компонен- | Уитни | почти одинакова; | |
| тов Р1,N1 | для N2, Р3 конкор- | |||
| P2, N2, Р3 | дантность МЗ выше | |||
| чем HP | ||||
| Дж.Полич | 10 пар МЗ | амплитуда | между клас- | для амплитуды Р3: |
| (J. Polich), | 20 пар HP | и латент- | совая | rМЗ = 0,64; |
| Т. Бенс | 18-30 | ный период | корреляция | rНР = -0,2; для ла- |
| (T.Burns), | Р3на | Пирсона | тентного периода | |
| редкие | Р3: rМЗ = 0,89; | |||
| тоны | rНР = -0,44 | |||
| Т. Роджерс | 10 пар МЗ | амплитуда | внутри- | для амплитуды Р3: |
| (Т. Rogers), | 10 пар ДЗ | и латентный | классовая | rМЗ = 0,50; |
| И. Деари | 18-60 | период Р3 | корреляция | rДЗ=0,35; |
| (I. Deary), | на редкие | для латентного | ||
| тоны | периода Р3: | |||
| rМЗ = 0,63; rДЗ = -0,2 | ||||
| С. О'Коннер | 59 пар МЗ | амплитуда | генетичес- | для амплитуды Р3: |
| с соавт. | 39 пар ДЗ | и латент- | кие модели | наследуемость от |
| (S. О'Соnnеr | 22-46 | ный период | 41 до 60%; латент- | |
| et al.), | Р3на | ный период не | ||
| редкие | наследуем | |||
| тоны |
Начиная с 60-х годов для изучения тонких нейрофизиологических механизмов избирательного внимания широко используют особый вариант эксперимента (oddball paradigm), в котором проводится сравнение ВП на два вида звуковых стимулов, различающихся по частоте тона и вероятности появления стимула [72, 134, 288]. Звуковые тоны поступают через наушники в левое или правое ухо, а испытуемому предлагается реагировать (нажимать на кнопку) на редко встречающиеся (целевые) стимулы и игнорировать часто встречающиеся (не-
редко сти-
целевые). В ответах на встречающиеся (целевые)
мулы возрастает амплитуда раннего компонента N1 (в интервале 80-180 мс) и существенно возрастает позднее позитивное
колебание — волна Р3 или Р300 (в интервале 250-600 мс). По современным представлениям,
эти компоненты отражают процессы переработки сенсорной информации и принятия решения при мобилизации селективного внимания (см. рис. 14.1). Причем, их амплитуды и латен-тности индивидуально специфичны и достаточно стабильны.
Известны четыре генетичес- Рис. 14.5. Распределение по коре боль-
ких исследования слуховых ВП ших полушарий статистически значи-при выполнении подобного за- мо наследуемых показателей ВП. дания (табл. 14.3). Они не рав- Локализация круга указывает отведения, v в - для которых :rМЗ отличалась от 0 с вероят-
ноценны по составу испытуе- ностью Р < 0,003; rМЗМЗ> rДЗ с вероятностью мых и способах оценки сходства Р < 0,01
близнецов, и это надо учиты- Диаметр круга пропорционален коэффи-вать при сопоставлении резуль- циенту наследуемости Н2. Масштаб —
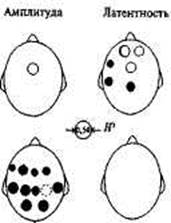
татов. Тем
той или
тельствуют
на параметры
регистрируемых
левые стимулы.
Наиболее из них О'Коннера
не менее все они
в
в центре рисунка. Аддитивный компо-
нент— пустой круг, доминантный ком-
иной степени свиде- понент — заполненный круг. Круг, наме-о влиянии генотипа ченный пунктиром, указывает отведение компонентов ВП, Р& исключенное из анализа
в ответ на це- а — Д™ компонента N1 в ответах на несигнальные стимулы; б — для компонента
Р3 в ответах на сигнальные стимулы представительным
является исследование с соавторами [346].
Оно было проведено с привлечением большой выборки близнецов, авторы использовали при анализе ВП современный метод картирования и генетический метод подбора моделей. При анализе ВП на нецелевые стимулы была установлена генотипическая обусловленность латентного периода экзогенного компонента N1, в лобно-ви-сочных отделах левого полушария (рис. 14.5). Оценки наследуемости находятся в пределах 0,43-0,63 и обнаруживают вклад как аддитивных, так и доминантных компонентов. Оценка наследуемости амплитуды N1 на эти же стимулы была близка к уровню достоверности и достигала его в отведении Cz (H = 0,60). Таким образом, изменчивость амплитудно-временных параметров экзогенного компонента N1
в ответах на нецелевые (часто встречающиеся) стимулы в значительной степени формируется под влиянием генотипа.
При оценке параметров эндогенного компонента Р300 в ответах на целевые (редко встречающиеся) стимулы были получены отчетливые доказательства наследственной обусловленности амплитуд этого компонента преимущественно в задних отделах коры больших полушарий. Генетический анализ показал, что наследуемость колеблется в пределах от 0,41 до 0,60. При этом влияния наследственных факторов на латентный период Р300 установлено не было (рис. 14.5).
Таким образом, природа межиндивидуальной вариативности амплитудных и временных параметров ВП на сенсорные стимулы (вспышки и тоны) меняется не только от интенсивности или содержательных особенностей стимула, но и в зависимости от условий задачи. Сосредоточение внимания" на стимуле и необходимость принятия решения (при выделении целевых стимулов) в целом приводят к увеличению доли генетической дисперсии в параметрах ВП.
Дата добавления: 2015-07-18; просмотров: 952;