Клубеньки у растений, не относящихся к бобовым
Корневые клубеньки или образования, напоминающие клубеньки, широко распространены на корнях не только бобовых растений. Они обнаружены у голосеменных и покрытосеменных двудольных растений.
Имеется до 200 видов различных растений, связывающих азот в симбиозе с микроорганизмами, образующими клубеньки на их корнях (или листьях).
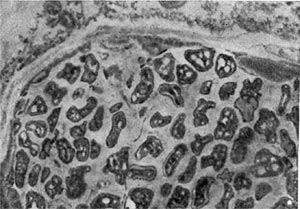
Рис. 168. Клубеньковая ткань ольхи (по Дж. Бекингу).
Клубеньки голосеменных растений (порядки Cycadales — саговники, Ginkgoales — гинкго-вые, Coniferales — хвойные) имеют ветвящуюся коралловидную, сферическую или четковидную форму. Они представляют собой утолщенные, видоизмененные боковые корни. Природа возбудителя, вызывающего их образование, до сих пор не выяснена. Эндофиты голосеменных растений относят и к грибам (фикомицетам), и к актиномицетам, и к бактериям, и к водорослям. Некоторые исследователи предполагают существование множественного симбиоза. Например, считают, что у саговников в симбиозе принимают участие азотобактер, клубеньковые бактерии и водоросли. Также не решен вопрос и о функции клубеньков у голосеменных. Ряд ученых пытается в первую очередь обосновать роль клубеньков как азотфиксаторов. Некоторые исследователи рассматривают клубеньки подокарповых как резервуары воды, а клубенькам саговников нередко приписывают функции воздушных корней.

Рис. 169. Клубеньки на корнях трибулюс (по Е. и О. Аллен).
У ряда представителей покрытосеменных двудольных растений клубеньки на корнях были обнаружены свыше 100 лет назад.
Сначала остановимся на характеристике клубеньков деревьев, кустарников и полукустарников (семейства Coriariaceae, Myricaceae, Be-tulaceae, Casuarinaceae, Elaeagnaceae, Rham-naceae), входящих в эту группу. Клубеньки большинства представителей данной группы — коралловидные гроздья розово-красного цвета, с возрастом приобретающие коричневую окраску. Имеются данные о наличии в них гемоглобина. У видов рода Elaeagnus (лоховые) клубеньки белого цвета.
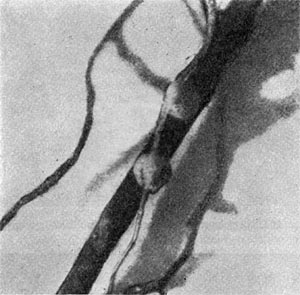
Рис. 170. Клубеньки на корнях вейштка лесного (по И.Л. Клевенской).
Нередко клубеньки имеют большие размеры. У казуарин (Casuarina) они достигают в длину 15 см. Функционируют несколько лет.
Растения с клубеньками распространены в разных климатических поясах или приурочены к определенному ареалу. Так, Shepherdia и Ceanothus встречаются только в Северной Америке, Casuarina — преимущественно в Австралии. Значительно шире распространены лоховые и облепиха.
Многие растения рассматриваемой группы произрастают на бедных питательными элементами почвах — песках, дюнах, скальных породах, болотах.
Подробнее всего изучены клубеньки ольхи (Alnus), в частности A. glutinosa, обнаруженные еще в 70-х годах прошлого столетия М. С. Ворониным (рис. 167). Существует предположение, что клубеньки свойственны не только современным, но и вымершим видам ольхи, поскольку их находили на корнях ископаемой ольхи в третичных отложениях долины реки Алдана — в Якутии.
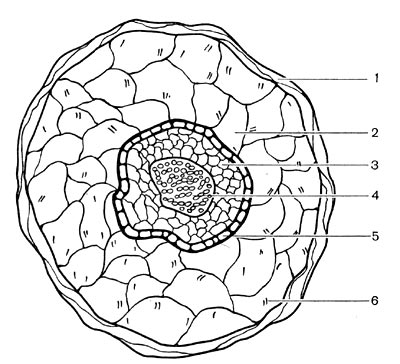
Рис. 171. Схема строения клубенька вейника лесного: 1 - кора, 2 — коровая паренхима; з — перициклическая паренхима; 4 - сосудистый пучок; 5 — эндодерма; 6 — бактерии (по И. Л. Клевенской).
Эндофит в клубеньках полиморфен. Он обычно встречается в виде гиф, везикул и бактероидов (рис. 168). Таксономическое положение эндофита до сих пор не установлено, поскольку многочисленные попытки выделить его в чистую культуру оказывались бесплодными, а если и удавалось выделить культуры, они оказывались невирулентными.
Основное значение всей этой группы растений, по-видимому, заключается в способности фиксировать молекулярный азот в симбиозе с эндофитом. Произрастая в областях, где разведение сельскохозяйственных растений с точки зрения экономики нерационально, они играют роль пионеров в освоении земли. Так, ежегодная прибавка азота в почве дюн Ирландии (мыс Верде) под посадками Casuarina equi-setifolia достигает 140 кг/га. Содержание азота в почве под ольхой на 30—50% выше, чем под березой, сосной, ивой. В высушенных листьях ольхи азота вдвое больше, чем в листьях других древесных растений. По расчетам А. Виртанена (1962), роща ольхи (в среднем 5 растений на 1 м2) дает за 7 лет прибавку азота 700 кг/га.
Значительно реже клубеньки встречаются у представителей семейства Zygophyllaceae (парнолистниковые). Впервые они были обнаружены Б. Л. Исаченко (1913) на корневой системе Tribulus terrestris. Позднее клубеньки были найдены и у других видов якор-цев.
Большинство представителей семейства Zygophyllaceae — ксерофитные кустарники или многолетние травы. Они распространены в пустынях тропических и субтропических областей, растут и на песчаных дюнах, пустошах и болотах умеренного пояса.
Интересно отметить, что такие тропические растения, как парнолистник ярко-красный, образуют клубеньки только при высокой температуре и низкой влажности почвы. Увлажнение почвы до 80% от полной влагоемкости препятствует формированию клубеньков. Как известно, у бобовых растений умеренного климата наблюдается обратное явление. При недостаточной влажности они не образуют клубеньков.
Клубеньки у растений семейства парнолист-никовых различаются размерами и расположением на корневой системе. Крупные клубеньки обычно развиваются на главном корне и близко от поверхности почвы. Более мелкие находятся на боковых корнях и на большей глубине. Иногда клубеньки образуются на стеблях, если они лежат на поверхности почвы.
Клубеньки у якорцев наземных на песках вдоль Южного Буга имеют вид мелких белых слегка заостренных или круглых бородавок.
Они обычно покрыты сплетением грибных гиф, проникающих внутрь коры корня.
У парнолистника ярко-красного клубеньки представляют собой концевые утолщения боковых корней растений. В клубеньках обнаруживаются бактероиды; бактерии очень напоминают клубеньковые.
Клубеньки тропических растений Tribulus cistoides твердые, округлые, около 1 мм в диаметре, соединены с корнями широким основанием, на старых корнях нередко мутовчатые. Чаще располагаются на корнях, чередуясь, с одной или с двух сторон (рис. 169). Для клубеньков характерно отсутствие зоны меристемы. Подобное явление отмечается при образовании клубеньков у хвойных растений. Клубенек поэтому возникает за счет деления клеток перицикла стелы.
Гистологическое изучение клубеньков Tribulus cistoides на разных стадиях развития показало, что в них отсутствуют микроорганизмы. На основании этого, а также скопления в клубеньках больших количеств крахмала их считают образованиями, выполняющими функцию обеспечения растений запасными питательными веществами.
Клубеньки вейника лесного — сферические или несколько удлиненные образования до 4 мм в диаметре, плотно сидящие на корнях растений (рис. 170). Цвет молодых клубеньков чаще всего белый, изредка розоватый, старых -желтый и бурый. Клубенек связан с центральным цилиндром корпя широким сосудистым пучком. Так же как и у Tribulus cistoides, клубеньки вейника имеют кору, коровую паренхиму, эндодерму, перициклическую паренхиму и сосудистый пучок (рис. 171).
Бактерии в клубеньках вейника лесного очень напоминают клубеньковых бактерий бобовых растений.
Клубеньки найдены на корнях капусты и редьки — представителей семейства крестоцветных. Предполагается, что их образуют бактерии, которые обладают способностью связывать молекулярный азот.
Среди растений семейства мареновых клубеньки встречаются у кофейных Coffea robusta и Goffea klainii. Они дихотомически ветвятся, иногда уплощены и имеют вид опахала. В тканях клубенька встречаются бактерии и бакте-роидные клетки. Бактерии, по мнению Стейар-та (1932), относятся к Rhizobium, но названы им Bacillus coffeicola.
Клубеньки у растений семейства розанных были обнаружены на дриаде (куропаточьей траве). У двух других представителей этого семейства — Purshia tridentata и Cercocarpus betuloides — описаны типичные коралловидные клубеньки. Однако никаких данных о строении этих клубеньков и природе их возбудителя в литературе пет.
Из семейства вересковых можно упомянуть только одно растение — медвежье ушко (или толокнянка), имеющее клубеньки на корневой системе. Многие авторы считают, что это коралловидные эктотрофные микоризы.
У покрытосеменных однодольных растений клубеньки распространены среди представителей семейства злаковых: лисохвоста лугового, мятлика лугового, волоснеца сибирского и волоснеца солончакового. Клубеньки образуются на концах корней; бывают продолговатыми, округлыми, веретеновидными. У лисохвоста молодые клубеньки светлые, прозрачные или полупрозрачные, с возрастом приобретают бурую или черную окраску. Данные о наличии бактерий в клетках клубеньков разноречивы.
Листовые клубеньки. Известно свыше 400 видов различных растений, образующих клубеньки на листьях. Наиболее хорошо изучены клубеньки у Pavetta и Psychotria. Они располагаются на нижней поверхности листьев вдоль основной жилки или рассеяны между боковыми жилками, имеют интенсивный зеленый цвет. В клубеньках сконцентрированы хлоропласты и танин. При старении на клубеньках часто появляются трещины.
Сформировавшийся клубенек заполнен бактериями, инфицирующими листья растения, очевидно, в момент прорастания семян. При выращивании стерильных семян клубеньки не возникают и растения развиваются хлоро-тичными. Выделенные из листовых клубеньков Psychotria bacteriophyla бактерии оказались принадлежащими к роду Klebsiella (К. rubiacearum). Бактерии фиксируют азот не только в симбиозе, но и в чистой культуре — до 25 мг азота на 1 г использованного сахара. Надо полагать, что они играют немаловажную роль в азотном питании растений на малоплодородных почвах. Есть основания полагать, что они снабжают растения не только азотом, но и биологически активными веществами.
Иногда на поверхности листьев можно увидеть глянцевые пленки или разноцветные пятна. Их образуют микроорганизмы филлосферы — особая разновидность эпифитных микроорганизмов, которые также участвуют в азотном питании растений. Бактерии филлосферы преимущественно олигонитрофилы (живут за счет ничтожных примесей азотсодержащих соединений в среде и, как правило, фиксируют небольшие количества молекулярного азота), тесно контактирующие с растением.
Свободноживущие азотфиксирующие микроорганизма. Азотобактер (AZOTOBACTER)
В 1901 году Бейеринк выделил из почвы аэробную неспорообразующую грамотрица-тельную бактерию, фиксирующую молекулярный азот, и назвал ее Azotobacter chroococcum (в родовом названии отражена способность бактерии фиксировать азот, в видовом — способность синтезировать коричневый пигмент — chroo и образовывать кокковидные клетки -coccum). Азотобактер — типичный представитель свободноживущих микроорганизмов. Свободноживущие — это все те микроорганизмы, которые живут в почве независимо от того, развивается вблизи растение или нет.
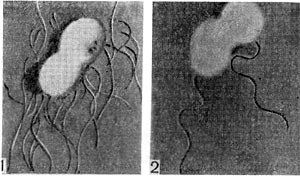
Рис. 173. Делящиеся клетки азотобактера (A. agilis), видны перитрихиалыше жгутики (7), у A. macrocyto-genes видны полярные жгутики (?) (по А. Бейлю и др.)-
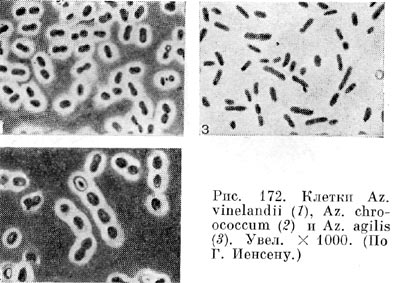
Культуры азотобактера в лабораторных условиях отличаются полиморфизмом. Клетки разных видов азотобактера в молодом возрасте показаны на рисунке 172. Молодые клетки азотобактера подвижны; они имеют многочисленные или единичные жгутики (рис. 173). У азотобактера обнаружены выросты, подобные фимбриям (рис. 174). В старых культурах клетки азотобактера покрываются плотной оболочкой, образуя цисты. Они могут прорастать, давая начало молодым клеткам (рис. 175).
Полиморфизм азотобактера зависит в значительной степени от состава среды, на которой он выращивается. На среде с этиловым спиртом (в качестве единственного источника углерода) азотобактер длительное время сохраняет подвижность и форму палочек. В то же время на многих других средах полиморфизм проявляется очень резко.
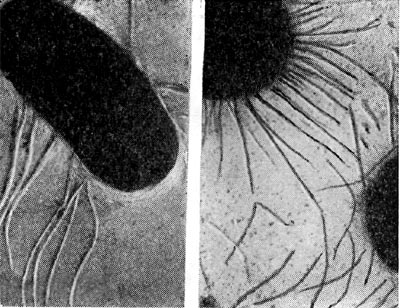
Рис. 174. Фимбрпоподобные образования у клеток азотобактера. Увел. X 24 000. (По Э. В. Болтянской.)
На плотных питательных средах, не содержащих азота, азотобактер образует крупные слизистые, иногда морщинистые колонии (рис. 176), окрашивающиеся при старении в желтовато-зеленоватый, розовый или коричнево-черный цвет. Колониям разных видов азотобактера присуща своя специфическая пигментация.
К настоящему времени известен ряд видов азотобактера: Azotobacter chroococcum, Az. beijerinckii, Az. vinelandii, Az. agilis, Az. nigricans, Az. galophilum.
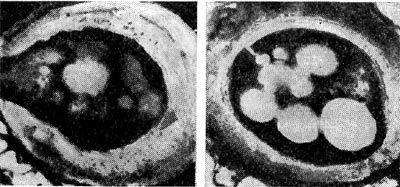
Рис. 175. Цисты азотобактера (по И. Чану и др.). Зрелая циста, наполненная гранулами жира и окруженная толстой плотной оболочкой (справа), и прорастающая циста (растущая молодая клетка разрывает оболочку цисты — слева). Увел. X 35 000.
Источником азота для азотобактера могут служить разнообразные минеральные (соли аммония, азотной и азотистой кислот) и органические (мочевина, различные аминокислоты) соединения. Однако если азотобактер развивается только за счет связанного в среде азота, он не выполняет своей основной функции — фиксации молекулярного азота. Азотобактер обычно фиксирует до 10—15 мг молекулярного азота на 1 г использованного источника углерода (например, глюкозы, сахарозы). Эта величина сильно колеблется в зависимости от условий выращивания культуры, состава питательной среды, ее кислотности, температуры, аэрации.
По отношению к источникам углерода В. Л. Омелянский (1923) назвал азотобактер полифагом («всеядным»).
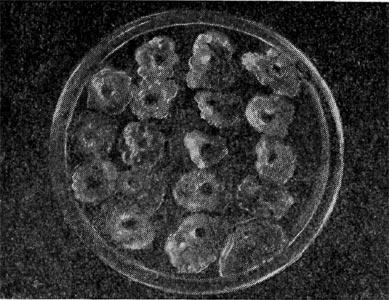
Рис. 176. Развитие колоний A. chroococcum вокруг комочков почвы па безазотной среде.
Азотобактер хорошо усваивает разнообразные углеводы (моно- и дисахара, некоторые полисахариды), органические кислоты, многоатомные спирты (глицерин, маннит) и другие вещества.
Многим исследователям удавалось выращивать азотобактер в чашках с питательной средой без азота и углерода, если чашки были помещены в камеру, где находились пары ацетона, этилового спирта или некоторых других органических соединений. В присутствии легкодоступных форм углеродсодержащих соединений азотобактер может частично использовать углекислоту из атмосферы. Повышение концентрации углекислого газа до 0,5% в воздухе несколько стимулирует развитие азотобактера. Но азотобактером лучше усваиваются легкодоступные формы углеродсодержащих органических соединений. В почве запас подвижного органического вещества невелик, поэтому именно недостаток легкодоступных соединений углерода в первую очередь ограничивает развитие азотобактера в природных условиях.
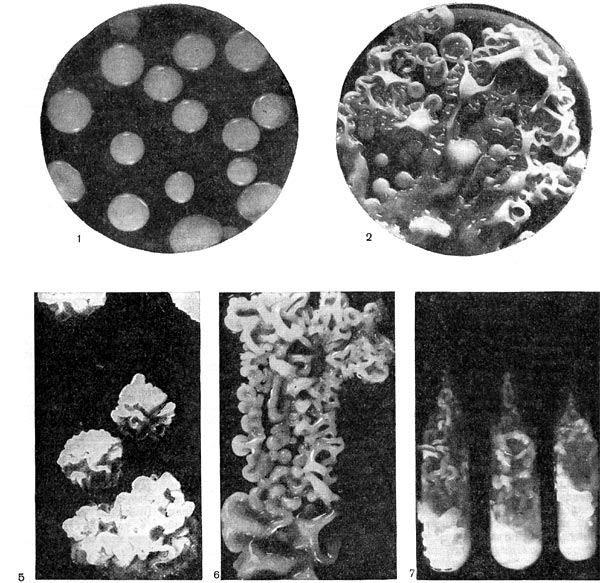
Рис. 177. Колонии бейеринкии разных типов (гладкие и складчатые варианты): 1—4, 6—8, 10 — по Н. И. Гогорикидзе; 5, 9, 11 — по Дж. Бекингу.
Какие же органические соединения может использовать в почве азотобактер? Перегнойные вещества почвы азотобактером практически не усваиваются. Поэтому в почвах, даже очень богатых перегноем, при отсутствии свежих органических остатков интенсивного размножения азотобактера не происходит.
Однако, если в почве имеются органические соединения и продукты распада растительных и животных клеток, азотобактер развивается хорошо. В частности, он усиленно размножается в почвах, удобренных соломой и соломистым навозом, а также в разнообразных компостах, содержащих целлюлозу. Азотобактер хорошо ассимилирует вещества, образующиеся при распаде целлюлозы.
Развитие азотобактера и фиксация им азота в значительной степени зависят от наличия в среде фосфора. Источником фосфора могут служить как органические, так и минеральные фосфорсодержащие соединения. Высокая чувствительность азотобактера к фосфору позволила разработать микробиологический метод определения потребности почв в фосфорных удобрениях.
В качестве тест-организма в этом методе используется азотобактер. Микробиологические методы определения потребности почвы в удобрении имеют ряд преимуществ перед химическими анализами, хотя, безусловно, уступают в точности.
Важную роль в обмене веществ азотобактера играет кальций. Этот элемент необходим азотобактеру при питании как молекулярным, так и аммонийным азотом (Г. Н. Зайцева, 1965). Недостаток кальция в среде приводит к сильной вакуолизации клеток и их вздутию.
Высокая чувствительность азотобактера к кальцию, так же как и к фосфору, используется для определения потребности почв в известковании.
Микроэлементы (молибден, бор, ванадий, железо, марганец) необходимы азотобактеру в первую очередь для осуществления процесса азотфиксации. Потребность в микроэлементах определяется в значительной степени геохимическими условиями существования азотобактера в почвах. Штаммы микроорганизма, выделенные из почв с высоким естественным содержанием того или иного микроэлемента, нуждаются, как правило, в более высоких концентрациях этих элементов.
Интересно, что радиоактивные элементы (радий, торий, уран) оказывают стимулирующий эффект на развитие азотобактера и процесс азотфиксации.
Азотобактер чрезвычайно чувствителен к реакции среды. Оптимальная для его развития область рН 7,2—8.2. Однако азотобактер способен развиваться и на средах с рН от 4,5 до 9,0; кислая реакция среды неблагоприятно действует на его развитие. Из кислых почв выделяются неактивные формы азотобактера, утратившие способность связывать молекулярный азот.
Большое влияние на развитие азотобактера оказывает влажность почвы. Клетки азотобактера имеют меньшее осмотическое давление, чем клетки грибов и актиномицетов; потребность во влаге аналогична потребности высших растений. Азотобактер распространен в пресных водоемах, илах, затопляемых рисовых полях, сточных водах, сильно увлажненных почвах, на водных растениях в прудах и водохранилищах. Это свидетельствует о его высокой степени гидрофильности. На основании высокой потребности во влаге почвенных форм азотобактера предполагается, что предки некоторых морских и почвенных видов азотобактера могли быть общими.
В отношении температуры азотобактер является типичным мезофильным организмом, с оптимумом развития около 25—30 °С. Понижение температуры азотобактер переносит хорошо, поэтому зимой даже в северных широтах численность его клеток в почве заметно не уменьшается.
Из биологических факторов, влияющих на развитие азотобактера, следует прежде всего отметить почвенные микроорганизмы. Они могут оказывать влияние на жизнедеятельность азотобактера в почве косвенно, изменяя, например, рН или окислительно-восстановительные условия, и непосредственно, вырабатывая питательные и биологически активные вещества. Так, активирующее влияние целлюлозоразру-шающих и маслянокислых микроорганизмов на развитие азотобактера и его антагонистические отношения с представителями почвенной микрофлоры отмечали многие советские и зарубежные исследователи. Биоценоз микроорганизмов, формирующийся в условиях той или иной почвы, меняется в значительной степени под влиянием растительного покрова. И азотобактер как член биоценоза также зависит от этого фактора. С помощью метода радиоавтографии установлено, что при нанесении меченных по фосфору клеток азотобактера на семена зерновых культур клетки обычно концентрируются вокруг растущей корневой системы проростков.
Имеются, однако, данные, что клеток азотобактера в ризосфере растений очень мало. В самом лучшем случае (при полном отсутствии антагонистов и благоприятных окружающих условиях) их количество не превышает 1 % от общего числа ризосферной микрофлоры.
Культуры азотобактера, как правило, образуют значительное количество биологически активных веществ: витамины группы В, никотиновую и пантотеновую кислоты, биотин, гетероауксин и гиббереллин. Однако, несмотря на то что культуры азотобактера вырабатывают целую серию биологически активных веществ, внесение витаминов, гиббереллина и гетероауксина в среду ускоряет рост азотобактера. Реакция на дополнительное внесение витаминов в среду является индивидуальной особенностью штаммов.
Азотобактер может продуцировать ростовые вещества типа ауксинов. Это подтверждается опытами, в которых было установлено образование дополнительных корешков у черенков фасоли под влиянием ауксинов, вырабатываемых азотобактером. Биологический тест — карликовая форма гороха сорта Пионер — позволяет определить в культуре азотобактера гиббереллиноподобные соединения.
Все эти соединения в совокупности способны стимулировать прорастание семян растений и ускорять их рост в тех, конечно, случаях, когда на корневой системе растений находится достаточное количество клеток азотобактера.
Кроме того, была обнаружена антагонистическая активность азотобактера по отношению к возбудителям бактериальных болезней растений. Азотобактер синтезирует фунгистати-ческий (задерживающий развитие грибов) антибиотик группы анисомицина. Ряд грибных организмов, встречающихся на семенах и в почве (виды из родов Fusarium, Alternaria, Penicillium), может угнетать развитие многих видов растений, особенно в холодную погоду. Азотобактер, продуцируя противогрибные антибиотические вещества, помогает растениям ра и развиваться, что имеет особенно болы значение в ранние фазы развития.
К сожалению, способность азотобактера тивно размножаться в почве и проявлять с: многогранные качества весьма ограничена за дефицита легкодоступных органических ществ в почве и высокой требовательности мироорганизма к окружающим условиям. Поэте стимулирующий эффект азотобактера проявляется лишь на плодородных почвах.
Распространение азотобактера в почвах Советского Союза имеет определенные закономерности. В целинных подзолах и дерново-подзолистых почвах, характеризующихся кислой реакцией, условия для развития азотобактера неблагоприятны. Только окультуривание таких почв создает возможности для его развития. В почвах с повышенным увлажнением и преобладанием луговой растительности (почвы пойм) азотобактер обычно встречается в течение всего вегетационного периода в больших количествах. В торфяниках азотобактер или отсутствует, или развивается очень слабо. В зоне достаточно увлажненных северных мощных черноземов азотобактер развивается хорошо, а в зоне обычных и южных черноземов при отсутствии орошения, а также в целинных и неполивных окультуренных каштановых почвах только как весенний эфемер. Максимальное развитие азотобактера в весенний период наблюдается и в целинных и в богарных почвах сероземной зоны. В солонцах и солончаках распространены преимущественно солестойкие расы азотобактера. В основном в почвах нашей страны доминирует Az. chroococcum.
Бейеринкия (BEIJERINCKIA)
Впервые аэробные бактерии рода Beijerinckia были выделены из кислых почв рисовых полей в Индии (в 1939 г.). Г. Деркс (1950), обнаружив эту бактерию в почве Ботанического сада в Богоре (Ява), предложил назвать ее именем М. Бейеринка — одного из первых исследователей фиксаторов азота.
Клетки бактерий рода Beijerinckia имеют круглую, овальную или палочковидную формы; палочки иногда искривлены. Размеры молодых клеток 0,5—2,0 X 1,0—4,5 мкм. Встречаются подвижные и неподвижные формы. Цист и спор не образуют. Культуры характеризуются медленным ростом. Типичные колонии формируются обычно через 3 недели при 30 °С. Большинство культур Beijerinckia образует на безазотном агаре с глюкозой выпуклые, нередко складчатые, блестящие слизистые колонии очень вязкой консистенции (рис. 177). При старении культуры, как правило, образуют темноокрашенный пигмент.
На 1 г использованного энергетического материала организмы рода Beijerinckia фиксируют 16—20 мг молекулярного азота. Спектр углеродсодержащих соединений, доступных бейеринкии, значительно уже, чем у азотобактера. Хорошо используются моно- и дисаха-риды, хуже — крахмал, органические кислоты, ароматические вещества не усваиваются. Минеральный азот и многие аминокислоты бактерии рода Beijerinckia предпочитают молекулярному азоту.
Основные отличия бейеринкии от азотобактера— высокая кислотоустойчивость(могут расти даже при рН 3,0), кальцефобность (ничтожные дозы кальция подавляют рост), устойчивость к высоким концентрациям железа, алюминия.
Бактерии рода Beijerinckia широко распространены в почвах южной и тропической зоны, реже встречаются в зоне умеренного климата. Beijerinckia часто встречается на поверхности листьев тропических растений в Индонезии.
Раньше полагали, что бактерии рода Beijerinckia могут существовать только в кислых почвах. Сейчас установлено, что они неплохо развиваются и в нейтральных и щелочных почвах. Тем не менее следует полагать, что Beijerinckia играют значительную роль в азотном балансе главным образом кислых почв (латеритах, красноземах), не имея существенного агрономического значения для нейтральных почв.
Клостридиум (CLOSTRIDIUM)
Первый анаэробный микроорганизм, усваивающий молекулярный азот, был выделен и описан С. Н. Виноградским в 1893 г. Он оказался спорообразующей бактерией, которой было дано наименование Clostridium pasteurianum (родовое название происходит от латинского слова clostrum — веретено; видовое — pasteurianum — дано в честь Луи Пастера).
Клетки Cl. pasteurianum крупные, их длина 2,5—7,5 мкм, ширина 0,7 —1,3 мкм. Располагаются они поодиночке, парами или образуют короткие цепочки. Молодые клетки подвижны, имеют перитрихиально расположенные жгутики, плазма их гомогенна. При старении клетки плазма становится гранулированной, в ней накапливается гранулеза (вещество типа крахмала). В центре клетки или ближе к ее концу формируется спора, которая в поперечнике значительно шире, чем вегетативная клетка, и поэтому клетка в этот период приобретает форму веретена. Размер спор 1,3 х 1,6 мкм. На рисунке 178 изображены клетки Cl. pasteurianum со спорами. Морфология спор и поведение ядерного вещества в процессе спорообразования у клостридиев детально описаны на стр. 228 В. И. Дудой.
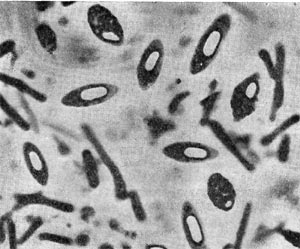
Рис. 178. Клетки Clostridium pasteurianum со спорами. Увел. X 3500 (по В.И. Дуде).
Азотфиксирующая функция выявлена у многих представителей рода Clostridium: Cl. pasteurianum, Cl. butyricum, Cl. butylicum, Cl. beijerinckia, Cl. pectinovorum, Cl. acetobutyli-cum и других видов. Наиболее энергичный азотонакопитель — Cl. pasteurianum — фиксирует 5—10 мг азота на 1 г потребленного источника углерода.
Наряду с молекулярным азотом бактерии рода Clostridium хорошо усваивают минеральные и органические азотсодержащие соединения. В качестве источника углеродного питания бактерии рода Clostridium используют различные соединения, которые обычно одновременно служат для них и источником энергии. К фосфору, калию и кальцию они значительно менее чувствительны, чем азотобактер. Однако удобрение почв фосфорно-калийными солями, известкование почв или компостов всегда приводит к возрастанию численности.
Клостридии относительно устойчивы к кислой и щелочной реакции среды. Область рН, при которой их развитие протекает нормально, довольно широка; минимальное значение рН ниже 4,5, максимальное — выше 8,5.
Влияние воздушно-водного режима на развитие бактерий рода Clostridium изучено достаточно полно. Будучи анаэробными, они хорошо переносят высокое насыщение почвы влагой. Однако оптимальная степень увлажнения для них определяется типом почвы и обеспеченностью органическим веществом. Лучше всего клостридий развивается при влажности почвы, равной 60—80% от полной вла-гоемкости.

Таблица 46. Бактериоз овса и суданской травы: 1 — бурый (красный) бактериоз овса; 2 — бактериоз суданской травы.
Больше всего бактерий рода Clostridium в верхних слоях почвы, которые богаты органическими веществами.
Бактерии рода Clostridium но-разному относятся к температуре, встречаются как мезофильные. так и термофильные бактерии. Молекулярный азот фиксируют лишь мезофилы.
У мезофильных форм оптимальная температура развития чаще всего находится в диапазоне 25—30 °С. Предел максимальных температур 37—45 °С.

Таблица 47. Болезни хлопчатника, табака и свеклы: 1 — гоммоз хлопчатника; 2 — бактериальная рябуха табака; 3 — серебристая болезнь свеклы (справа — пятна при увеличении).
Споры клостридиев весьма устойчивы к высоким температурам. Они выдерживают нагревание при 75 °С в течение 5 ч и в течение 1 ч нагревание при 80 °С. Споры термофильных клостридиев погибают при кипячении через 30 мцн. Более высокая температура (110 °С) быстро убивает их.
Со многими микроорганизмами в почве Clostridium находится в метабиотических отношениях, при которых предполагается обмен продуктами метаболизма. Так, азотобактер улучшает условия жизни клостридия, поглощая кислород, а клостридий вырабатывает из органических соединений, недоступных азотобактеру, органические кислоты, которые может ассимилировать азотобактер.
Было бы трудно ответить на вопрос: в каких почвах нет Clostridium? «Всеядность» Clostridium, малая взыскательность к условиям внешней среды, а также способность в неблагоприятных условиях переходить в состояние спор объясняют их широкое, практически повсеместное распространение.
Накопления азота в почвах за счет деятельности Clostridium, однако,невелики и не превышают, как правило, нескольких килограммов на один гектар почвы.
Дата добавления: 2015-06-12; просмотров: 1939;