Физиология
Большинство пурпурных серобактерий — строгие анаэробы и облигатные фототрофы, т. е. рост их возможен только при освещении. Известно лишь три вида, растущие в присутствии воздуха, причем не только на свету, но и в темноте, хотя и медленно. Это — A. roseus, Е. shaposhnikovii и Т. roseopersicina. Все несерные пурпурные бактерии также растут в анаэробных условиях, но в основном являются факультативными аэробами. До недавнего времени считали, что рост пурпурных бактерий в темноте возможен лишь в аэробных или микроаэрофильных условиях, так как в отсутствие света они получают энергию в процессе дыхания. Однако недавно установлено, что R. rubrum и ряд представителей Rhodopseudomonas растут в темноте и в строго анаэробных условиях за счет сбраживания некоторых органических субстратов. Такую же возможность, видимо, имеют пурпурные серобактерии Е. shaposhnikovii и Т. roseopersicina.
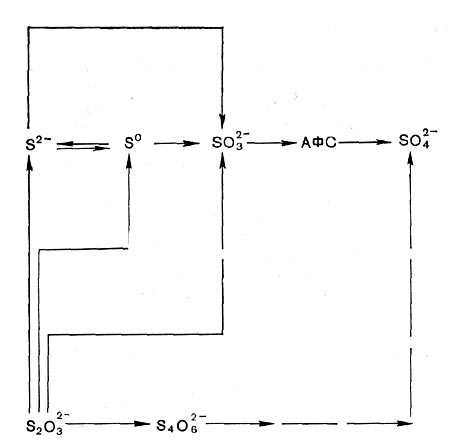
Рис. 128. Пути окисления соединений серы фототрофными бактериями АФС — аденилнлсульфат.
Зеленые бактерии — строгие анаэробы и облигатные фототрофы. Исключением являются представители рода Chloroflexis. Они растут только в аэробных условиях, причем и при освещении и в темноте. Однако даже фототроф-ные бактерии, хорошо растущие в темноте, лучше развиваются при наличии света. В зависимости от организма оптимальные условия освещения для его роста могут быть различны. Одни виды хорошо растут при слабом освещении (100—300 лк), другие — при более сильном свете (700—2000 лк).
Хотя неоднократно отмечалось развитие фо-тотрофных бактерий в горячих источниках с температурой воды 60—80 °С, ни одного обли-гатно термофильного вида до сих пор не выделено. Оптимальная температура для роста многих видов этих микроорганизмов в лабораторных условиях — 25—35 °С. Только Е. halo-phila имеет температурный оптимум около 47 °С. В то же время известны пурпурные и зеленые серобактерии (Thiopedia, Lamprocystis, Pelodictyon), хорошо растущие при температуре не выше 20 °С.
Фототрофные бактерии в целом могут расти в достаточно широком интервале значений рН, примерно от 5,0 до 11,0, хотя для отдельных видов и штаммов оптимальное значение рН и зона, в которой возможен их рост, могут существенно различаться. Для многих представителей пурпурных и зеленых бактерий оптимальное значение рН 7,0—7,5. Но известны виды, для которых оптимально значение рН 6,0—6,5. Для Rh. acidophila оно составляет даже 5,8. Напротив, некоторые штаммы Е. shaposhnikovii хорошо растут при рН 8,5 — 9,0. Показано также, что оптимальное значение рН для роста фототрофных бактерий может несколько меняться в зависимости от состава среды.
Все фототрофные бактерии растут на простых синтетических средах, что облегчает выявление их потребности в отдельных элементах. Обязательными компонентами сред, кроме источников углерода и азота, естественно, являются фосфор, сера, калий и магний в виде минеральных солей. Установлено также, что эти микроорганизмы нуждаются в довольно большом количестве железа, но по сравнению с растениями проявляют значительно меньшую потребность в марганце. Рост их зависит от концентрации кальция и наличия в небольших количествах таких элементов, как Mo, Co, Zn, Си и, видимо, других.
Выше отмечалось, что фототрофные бактерии встречаются как в пресных, так и в соленых водоемах, причем некоторые пурпурные бактерии растут при концентрации хлористого натрия больше 20%. Наиболее галофильной формой является Е. halophila. Оптимальная концентрация NaCl для этого организма 14—22%. Для других видов, даже выделенных из очень соленых водоемов, она более низкая. Зеленые бактерии растут при концентрации NaCl не, более 10—11%. И для пурпурных и зеленых бактерий, выделенных из соленых водоемов, присутствие NaCl обязательно. Так, морские штаммы растут обычно хорошо в среде, содержащей 1—2% NaCl.
За исключением отдельных мутантов, все фототрофные бактерии используют в качестве источника азота соли аммония. Способность к ассимиляционной нитратредукции проявляется довольно редко. Некоторые пурпурные бактерии, в первую очередь несерные, используют как источники азота мочевину и различные аминокислоты, а также растут на средах с пептоном. У многих пурпурных и зеленых бактерий установлена способность фиксировать молекулярный азот.
В качестве источника серы для синтеза серу-содержащих компонентов клеток несерные пурпурные бактерии и некоторые пурпурные серобактерии могут использовать сульфаты. Однако многие представители пурпурных серобактерий и зеленые серобактерии способностью к ассимиляционной сульфатредукции не обладают и могут использовать серу лишь в восстановленной форме, в виде сульфида, тиосульфата или цистеина.
Кроме того, пурпурные и зеленые серобактерии используют восстановленные соединения серы как доноры водорода (Н-доноры) при фотоассимиляции углекислоты. Все эти микроорганизмы способны окислять сероводород с образованием сульфатов, но сначала образуется молекулярная сера (рис. 128). У большинства пурпурных серобактерий сера откладывается в клетках. Зеленые серобактерии и представители рода Ectothiorhodospira накапливают серу в среде. Кроме сульфида и молекулярной серы, многие пурпурные и зеленые серобактерии окисляют тиосульфат, а некоторые виды тетра-тионат, сульфит и тиогликолят.
До последнего времени считали, что несерные пурпурные бактерии сероводород не окисляют и лишь для одного вида (Rh. palustris) была показана способность окислять тиосульфат. Однако недавно установлено, что если поддерживать низкую концентрацию сульфида в проточных условиях культивирования, то такие несерные пурпурные бактерии, как R. rubrum, Rh. capsulata, Rh. palustris, Rh. spheroides, окисляют его и растут. При этом Rh. palustris образует сульфаты. У остальных видов окисление сульфида заканчивается образованием молекулярной серы, которая обнаруживается в среде.
Эти данные весьма важны, поскольку до сих пор деление пурпурных бактерий на серные и несерные основывалось на их способности окислять сероводород. Сейчас такой принцип оказывается неприемлемым. Поэтому предлагается учитывать, что пурпурные серобактерии окисляют сероводород до сульфатов через образование молекулярной серы, а несерные пурпурные бактерии окисляют его только до серы или до сульфатов, но без накопления серы как промежуточного продукта. Не исключено, однако, что в дальнейшем эти критерии также нельзя будет использовать.
Кроме соединений серы, многие пурпурные и зеленые бактерии способны окислять молекулярный водород.
Большинство представителей несерных пурпурных бактерий нуждается в одном или нескольких витаминах группы В: тиамине, биотине, никотиновой, параминобензойной кислотах. Потребность в витаминах не выявлена лишь у Rhodomicrobium vannielii, Rh. acidophila и Rhodospirillum tenue. Среди пурпурных и зеленых серобактерий значительно больше организмов, которые не нуждаются в витаминах.
Однако есть виды и штаммы, требующие готового витамина В12. Такую потребность проявляют, например, крупные формы Chromatium (Chr. okenii, Chr. buderi и другие), а также Thiospirillum jenense.
Фотосинтезирующие бактерии делят на фотоавтотрофы и фотогетеротрофы. К первым принадлежит ряд пурпурных и зеленых серобактерий, а также отдельные представители несерных пурпурных бактерий, способные расти на чисто минеральных средах. Для таких микроорганизмов единственным источником углерода может служить углекислота, обычно вносимая в виде бикарбоната. К фотогетеротро-фам относят большинство несерных пурпурных бактерий, поскольку они растут лишь при наличии органических соединений. Строго говоря, к автотрофам нельзя причислять и фото-синтезирующих бактерий, которые нуждаются в готовых витаминах. С другой стороны, следует отметить, что фотогетеротрофные бактерии часто фиксируют в больших количествах углекислоту. В то же время все автотрофные представители этих микроорганизмов способны использовать готовые органические соединения, хотя возможности в этом отношении у разных видов неодинаковы.
Более разнообразные органические соединения могут использовать несерные пурпурные бактерии. К их числу относятся низшие жирные кислоты от С3 (муравьиная) до С9 (пеларгоновая), дикарбоновые кислоты, окси- и кетокислоты (от С3 до С6), такие, как пировиноградная, молочная, яблочная, янтарная и др.
Используются некоторые сахара (в частности, глюкоза, фруктоза, манноза), спирты (этанол, изопропанол, маннит, сорбит), соединения ароматического ряда (бензоат, пара-оксибензоат, манделат, катехин), а также другие циклические соединения. Однако спектр органических субстратов, обеспечивающих рост различных видов и штаммов этих микроорганизмов, неодинаков.
Пурпурные и особенно зеленые серобактерии, как правило, используют меньшее число органических соединений; чаще всего отдельные органические кислоты. У некоторых видов возможности исчерпываются потреблением ацетата и пирувата.
Для несерных и некоторых пурпурных серобактерий (Е. shaposhnikovii, Chr. vinosum) органические соединения могут служить источниками углерода и Н-донорами при фотоассимиляции углекислоты, а также в других восстановительных процессах. Ряд видов способен в темноте окислять органические соединения с получением энергии, обеспечивающей их рост. У зеленых и многих пурпурных серобактерий способности ограничиваются использованием органических соединений при фотосинтезе лишь как дополнительных (по отношению к С02) источников углерода.
Дата добавления: 2015-06-12; просмотров: 979;