Структура бактериальной клетки
Структура бактерий хорошо изучена с помощью электронной микроскопии целых клеток и их ультратонких срезов. Бактериальная клетка состоит из клеточной стенки, цитоплазматической мембраны, цитоплазмы с включениями и ядра, называемого нуклеоидом. Имеются дополнительные структуры: капсула, микрокапсула, слизь, жгутики, пили (рис.1); некоторые бактерии в неблагоприятных условиях способны образовывать споры.
Клеточная стенка — прочная, упругая структура, придающая бактерии определенную форму и вместе с подлежащей цитоплазматической мембраной «сдерживающая» высокое осмотическое давление в бактериальной клетке. Она участвует в процессе деления клетки и транспорте метаболитов. Наиболее толстая клеточная стенка у грамположительных бактерий (рис.1). Так, если толщина клеточной стенки грамотрицательных бактерий около 15—20 нм, то у грамположительных она может достигать 50 нм и более. В клеточной стенке грамположительных бактерий содержится небольшое количество полисахаридов, липидов, белков.
Основным компонентом клеточной стенки этих бактерий является многослойный пептидогликан (муреин, мукопептид), составляющий 40—90 % массы клеточной стенки.
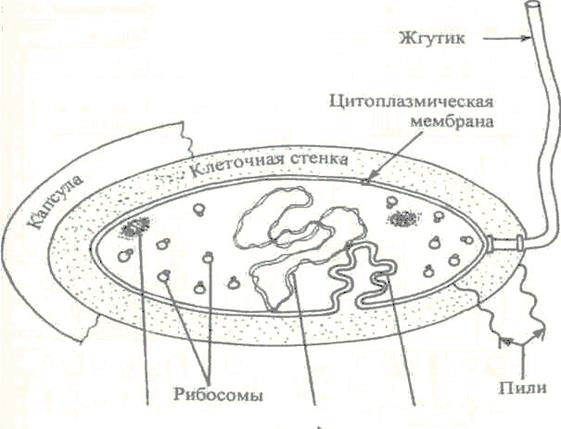
Волютин Мезосома Нуклеоид
Рис. 1. Строение бактериальной клетки.
С пептидогликаном клеточной стенки грамположительных бактерий ковалентно связаны тейхоевые кислоты (от греч. teichos — стенка), молекулы которых представляют собой цепи из 8—50 остатков глицерола и рибитола, соединенных фосфатными мостиками. Форму и прочность бактериям придает жесткая волокнистая структура многослойного с поперечными пептидными сшивками пептидогликана. Пептидогликан представлен параллельно расположенными молекулами гликана, состоящего из повторяющихся остатков N-ацетилглюкозамина и N-ацетилмурамовой кислоты, соединенных гликозидной связью типа Р (1 —> 4).
Лизоцим, являясь ацетилмурамидазой, разрывает эти связи. Гликановые молекулы связаны поперечной пептидной связью. Отсюда и название этого полимера — пептидогликан. Основу пептидной связи пептидогликана грамотрицательных бактерий составляют тетрапептиды, состоящие из чередующихся L- и Д-аминокислот.
У E. coli пептидные цепи соединены друг с другом через D- аланин одной цепи и мезодиаминопимелиновую кислоту другой.
Состав и строение пептидной части пептидогликана у грамотрицательных бактерий стабильны в отличие от пептидогликана грамположительных бактерий, аминокислоты которого могут отличаться по составу и последовательности. Тетрапептиды здесь соединены друг с другом полипептидными цепочками из 5 остатков глицина. У грамположительных бактерий вместо мезодиаминопимелиновой кислоты часто содержится лизин. Фосфолипид
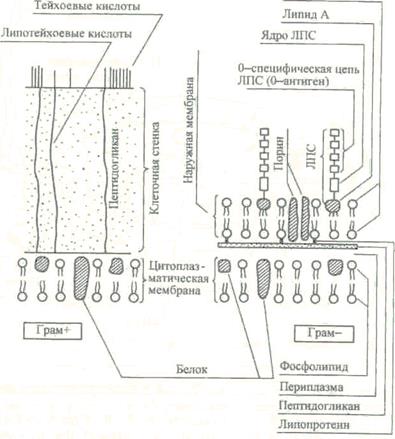
Рис. 2. Строение поверхностных структур грамположительных (грам+) и грамотрицательных (грам") бактерий.
Элементы гликана (ацетилглюкозамин и ацетилмурамовая кислота) и аминокислоты тетрапептида (мезодиаминопимелиновая и Л-глутаминовая кислоты, Д-аланин) являются отличительной особенностью бактерий, поскольку они и Д-изомеры аминокислот отсутствуют у животных и человека.
Способность грамположительных бактерий при окраске по Граму удерживать генциановый фиолетовый в комплексе с йодом (сине-фиолетовая окраска бактерий) связана со свойством многослойного пептидогликана взаимодействовать с красителем. Кроме этого, последующая обработка мазка бактерий спиртом вызывает суживание пор в пептидогликане и тем самым задержку красителя в клеточной стенке. Грамотрицательные бактерии после воздействия спиртом утрачивают краситель, обесцвечиваются и при обработке фуксином окрашиваются в красный цвет. Это обусловлено меньшим количеством пептидогликана (5—10 % массы клеточной стенки).
В состав клеточной стенки грамотрицательных бактерий входит наружная мембрана, связанная посредством липопротеина с подлежащим слоем пептидогликана (рис.2). Наружная мембрана представляет собой волнообразную трехслойную структуру, сходную с внутренней мембраной, которую называют ци-топлазматической. Основным компонентом этих мембран служит бимолекулярный (двойной) слой липидов.
Наружная мембрана является асимметричной мозаичной структурой, представленной липополисахаридами, фосфолипидами и белками. С ее внешней стороны расположен липополисахарид (ЛПС), состоящий из трех компонентов: липида А, стержневой части, или ядра (лат. core — ядро), и 0-специфической цепи полисахарида, образованной повторяющимися олигосахаридными последовательностями.
Липополисахарид «заякорен» в наружной мембране липидом А, обусловливающим токсичность ЛПС, отождествляемому поэтому с эндотоксином. Разрушение бактерий антибиотиками приводит к освобождению большого количества эндотоксина,чтоможет привести к эндотоксическому шоку больного.
От липида А отходит ядро, или стержневая часть ЛПС. Наиболее постоянной частью ядра ЛПС является кетодезоксиоктоновая кислота (3-деокси-г)-манно-2-октулосоновая кислота). 0-специфическая цепь, отходящая от стержневой части молекулы ЛПС, обусловливает серогруппу, серовар (разновидность бактерий, выявляемая с помощью иммунной сыворотки) определенного штамма бактерий. Таким образом, с понятием ЛПС — связаны представления об 0-антигене, покоторому можно дифференцировать бактерии. Генетические изменения могут привести к изменениям в биосинтезе компонентовЛПС бактерий и к появлению в результате этого L -форм.
Белки матрикса наружной мембраны пронизывают ее таким образом, что молекулы белка, называемые поринами, окаймляют гидрофильные поры, через которые проходят вода и мелкие молекулы с относительной массой до 700. Между наружной и цитоплазматической мембранами находится периплазматическое пространство, или периплазма, содержащая ферменты. При нарушении синтеза клеточной стенки бактерий под влиянием лизоцима, пенициллина, защитных факторов организма и других соединений образуются клетки с измененной (часто шаровидной) формой: протопласты — бактерии, полностью лишенные клеточной стенки; сферопласты — бактерии с частично сохранившейся клеточной стенкой. После удаления ингибитора клеточной стенки такие измененные бактерии могут реверсировать, т.е. приобретать полноценную клеточную стенку и восстанавливать исходную форму.
Бактерии сферо- или протопластного типа, утратившие способность к синтезу пептидогликана под влиянием антибиотиков или других факторов и способные размножаться, называются L-формами (от названия института им. Листера). L -формы могут возникать и в результате мутаций. Они представляют собой осмотически чувствительные, шаровидные, колбовидные клетки различной величины, в том числе и проходящие через бактериальные фильтры. Некоторые L-формы (нестабильные) при удалении фактора, приведшего к изменениям бактерий, могут реверсировать, «возвращаясь» в исходную бактериальную клетку. L -формы могут образовывать многие возбудители инфекционных болезней.
Цитоплазматическая мембрана при электронной микроскопии ультратонких срезов представляет собой трехслойную мембрану, окружающую наружную часть цитоплазмы бактерий. По структуре она похожа на плазмалемму клеток животных и состоит из двойного слоя липидов, главным образом фосфолипидов с внедренными поверхностными, а также интегральными белками, как бы пронизывающими насквозь структуру мембраны. Некоторые из них являются пермеазами, участвующими в транспорте веществ. Цитоплазматическая мембрана является динамической структурой с подвижными компонентами, поэтому ее представляют как мобильную текучую структуру. Она участвует в регуляции осмотического давления, транспорте веществ и энергетическом метаболизме клетки (за счет ферментов цепи переноса электронов, аденозинтрифосфатазы и др.). При избыточном росте (по сравнению с ростом клеточной стенки) Цитоплазматическая мембрана образует инвагинаты — впячивания в виде сложно закрученных мембранных структур, называемые мезосомами. Менее сложно закрученные структуры называются внутрицитоплазматическими мембранами. Роль мезосом и внутрицитоплазматических мембран до конца не выяснена. Предполагают даже, что они являются артефактом, возникающим после приготовления (фиксации) препарата для электронной микроскопии. Тем не менее считают, что производные цитоплазматической мембраны участвуют в делении клетки, обеспечивая энергией синтез клеточной стенки, принимают участие в секреции веществ, спорообразовании, т.е. в процессах с высокой затратой энергии.
Цитоплазмазанимает основной объем бактериальной клетки и состоит из растворимых белков, рибонуклеиновых кислот, включений и многочисленных мелких гранул — рибосом, ответственных за синтез (трансляцию) белков. Рибосомы бактерий имеют размер около 20 нм и коэффициент седиментации 70S, 3 отличие от 80^-рибосом, характерных для эукариотических клеток. Поэтому некоторые антибиотики, связываясь с рибосо-мами бактерий, подавляют синтез бактериального белка, не влияя на синтез белка эукариотических клеток. Рибосомы бактерий могут диссоциировать на две субъединицы — 50S и 30S. В цитоплазме имеются различные включения в виде гранул гликогена, полисахаридов, поли-р-масляной кислоты и полифосфатов (волютин). Они накапливаются при избытке питательных веществ в окружающей среде и выполняют роль запасных веществ для питания и энергетических потребностей. Волютин обладает сродством к основным красителям, обладает метахро-мазией и легко выявляется с помощью специальных методов окраски. Характерное расположение зерен волютина выявляется у дифтерийной палочки в виде интенсивно прокрашивающихся полюсов клетки.
Нуклеоид — эквивалент ядра у бактерий. Он расположен в центральной зоне бактерий в виде двунитчатой ДНК, замкнутой в кольцо и плотно уложенной наподобие клубка. В отличие от эукариот ядро бактерий не имеет ядерной оболочки, ядрышка и основных белков (гистонов). Обычно в бактериальной клетке содержится одна хромосома, представленная замкнутой в кольцо молекулой ДНК. При нарушении деления в ней может находиться 4 и более хромосом. Нуклеоид выявляется в световом микроскопе после окраски специфическими для ДНК методами: по Фельгену или по Романовскому—Гимзе. На электронограммах ультратонких срезов бактерий нуклеоид имеет вид светлых зон с фибриллярными, нитевидными структурами ДНК, связанной определенными участками с цитоплазматической мембраной или мезосомой, участвующими в репликации хромосомы.
Кроме нуклеоида, представленного одной хромосомой, в бактериальной клетке имеются внехромосомные факторы наследственности — плазмиды, представляющие собой ковалентно замкнутые кольца ДНК.
Капсула — слизистая структура толщиной более 0,2 мкм, прочно связанная с клеточной стенкой бактерий и имеющая четко очерченные внешние границы. Капсула различима в мазках-отпечатках из патологического материала. В чистых культурах бактерий капсула образуется реже. Она выявляется при специальных методах окраски по Бурри—Гинсу, создающих негативное контрастирование веществ капсулы.
Обычно капсула состоит из полисахаридов (экзополисахаридов), иногда из полипептидов, например у сибиреязвенной бациллы. Капсула гидрофильна, она препятствует фагоцитозу бактерий.
Многие бактерии образуют микрокапсулу — слизистое образование толщиной менее 0,2 мкм, выявляемое лишь при электронной микроскопии. От капсулы следует отличать слизь — мукоидные экзополисахариды, не имеющие четких внешних границ. Мукоидные экзополисахариды характерны для мукоидных штаммов синегнойной палочки, часто встречающихся в мокроте больных с кистозным фиброзом. Бактериальные экзополисахариды участвуют в адгезии (прилипании к субстратам), их еще называют гликокаликсом. Кроме синтеза экзополисахаридов бактериями, существует и другой механизм их образования: путем действия внеклеточных ферментов бактерий на дисахариды. В результате этого образуются декстраны и леваны. Капсула и слизь предохраняют бактерии от повреждений, высыхания, так как, являясь гидрофильными, хорошо связывают воду, препятствуют действию защитных факторов макроорганизма и бактериофагов.
Жгутики бактерий определяют подвижность бактериальной клетки. Жгутики представляют собой тонкие нити, берущие начало от цитоплазматической мембраны, имеют большую длину, чем сама клетка (рис.3). Толщина жгутиков 12—20 нм, длина 3—12 мкм. Число жгутиков у бактерий различных видов варьирует от одного (монотрих) у холерного вибриона до десятка и сотен жгутиков, отходящих по периметру бактерии (пери-трих) у кишечной палочки, протея и др. Лофотрихи имеют пучок жгутиков на одном из концов клетки. Амфитрихи имеют по одному жгутику или пучку жгутиков на противоположных концах клетки. Жгутики прикреплены к цитоплазматической мембране и клеточной стенке специальными дисками. Жгутики состоят из белка — флагеллина (от naT.flagellum — жгутик), обладающего антигенной специфичностью. Субъединицы флагеллина закруче ны в виде спирали. Жгутики выявляют с помощью электронной микроскопии препаратов, напыленных тяжелыми металлами, или в световом микроскопе после обработки специальными методами, основанными на протравливании и адсорбции различных веществ, приводящих к увеличению толщины жгутиков (например, после серебрения).
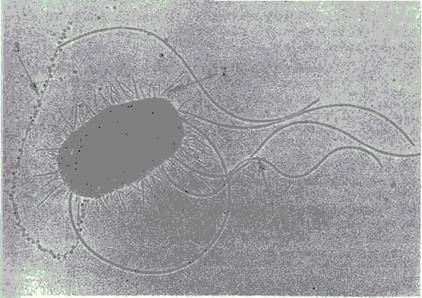
Рис. 3. Кишечная палочка. Электронограмма (препарат В.С.Тюрина). 1 — жгутики, 2 — ворсинки, 3 — F-пили.
Ворсинки, или пили (фимбрии), — нитевидные образования (рис.3), более тонкие и короткие (3—10 нм х 0,3—10 мкм), чем жгутики. Пили отходят от поверхности клетки и состоят из белка пилина. Они обладают антигенной активностью. Среди пилей выделяются: пили, ответственные за адгезию, т.е. за прикрепление бактерий к поражаемой клетке (пили 1-го типа, или общего типа — common pili), пили, ответственные за питание, водно-солевой обмен; половые (F-пили), или конъюга-ционные пили (пили 2-го типа). Пили общего типа многочисленны — несколько сотен на клетку. Половые пили образуются так называемыми «мужскими» клетками-донорами, содержащими трансмиссивные плазмиды (F, R, Col). Их обычно бывает 1—3 на клетку. Отличительной особенностью половых пилей является взаимодействие с особыми «мужскими» сферически ми бактериофагами, которые интенсивно адсорбируются на половых пилях.
Споры — своеобразная форма покоящихся фирмикутных бактерий, т.е. бактерий с грамположительным типом строения клеточной стенки.
Споры образуются при неблагоприятных условиях существования бактерий (высушивание, дефицит питательных веществ и др.). При этом внутри одной бактерии образуется одна спора. Образование спор способствует сохранению вида и не является способом размножения, как у грибов.
Спорообразующие аэробные бактерии, у которых размер споры не превышает диаметр клетки, иногда называются бациллами. Спорообразующие анаэробные бактерии, у которых размер споры превышает диаметр клетки, и поэтому они принимают форму веретена, называются клостридиями (лат. clostridium — веретено).
Процесс спорообразования (споруляция) проходит ряд стадий, в течение которых часть цитоплазмы и хромосома отделяются, окружаясь цито плазматической мембраной; образуется проспора, затем формируется многослойная плохо проницаемая оболочка. Спорообразование сопровождается интенсивным потреблением проспорой, а затем формирующейся оболочкой споры дипиколиновой кислоты и ионов кальция. После формирования всех структур спора приобретает термоустойчивость, которую связывают с наличием дипиколината кальция. Спорообразование, форма и расположение спор в клетке (вегетативной) являются видовым свойством бактерий, что позволяет отличать их друг от друга. Форма спор может быть овальной, шаровидной, расположение в клетке — терминальное, т.е. на конце палочки (возбудитель столбняка), субтерминальное — ближе к концу палочки (возбудители ботулизма, газовой гангрены) и центральное (сибиреязвенная бацилла).
Дата добавления: 2015-06-12; просмотров: 7251;