Отвечает на стимул
Как мы уже отмечали, с позиций парадигмы реактивности поведение индивида представляет собой реакцию на стимул. В основе реакции лежит проведение возбуждения по рефлекторной дуге: от рецепторов через центральные структуры к исполнительным органам.
Нейрон при этом оказывается элементом, входящим в рефлекторную дугу, а его функция — обеспечением проведения возбуждения. Тогда совершенно логично рассмотреть детерминацию активности этого элемента следующим образом: «...ответ на стимул, подействовавший на некоторую часть ее (нервной клетки. — Ю. А.) поверхности, может распространяться дальше по клетке и действовать как стимул на другие нервные клетки...» [Бринк, 1960, с. 93]. Следовательно, в рамках парадигмы реактивности рассмотрение нейрона вполне методологически последовательно: нейрон, как и организм, реагирует на стимулы. В качестве стимула выступает импульсация, которую нейрон получает от других клеток, в качестве реакции — следующая за синаптическим притоком импульсация данного нейрона (рис. 14.2).
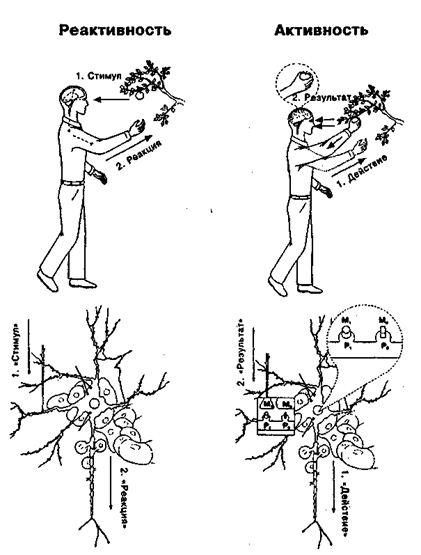
Рис. 14.2. Индивид и нейрон в парадигмах активности и реактивности
Цифрами на схеме обозначена последовательность событий. В соответствии с парадигмой реактивности за стимулом (1) следует реакция (2) — поведенческая у человека, импульсная у нейрона. В последнем случае в качестве стимула рассматривается импульсация нейрона, аксон которого (параллельный стрелке с надписью «Стимул») контактирует с дендритом реагирующего нейрона, окруженного глиальными клетками и соприкасающегося с кровеносным сосудом, который расположен над словом «Реакция». Реакция представляет собой импульсные разряды реагирующего нейрона. В соответствии с парадигмой активности действие (1) (поведенческое — у человека, импульсация — у нейрона) завершается достижением результата и его оценкой (2). Пунктирной линией обозначена модель будущего результата: для человека — контакт с объектом-целью, для нейрона — получение соответствующих метаболитов (М1 — от контактирующего нейрона; М2 — от соседней глиальной клетки), которые соединяются с рецепторами нейрона (Р1, Р2)
К сожалению, такая методологическая последовательность отсутствовала в рамках парадигмы активности. Как правило, анализ «нейронных механизмов» целенаправленного поведения приводил авторов к тому, что мы назвали ранее «уровневой эклектикой»: представлению о том, что индивид осуществляет целенаправленное поведение, а его отдельный элемент — нейрон — реагирует на приходящее к нему возбуждение — стимул. Важнейшей задачей стало устранение подобной эклектики.
Подход к нейрону как к проводнику возбуждения встречал возражения уже давно, например со стороны Дж. Э. Когхилла, который, однако, не мог в отсутствие целостной и последовательной теории, вписывающейся в парадигму активности, дать решение, адекватное сформулированной задаче. В его концепции нейрон реагирует «на окружающую среду так же, как живой организм» [Когхилл, 1934, с. 56]. Решающий шаг в направлении решения этой задачи был сделан П. К. Анохиным [1975], который в своей последней работе подверг аргументированной критике общепринятую, как он ее назвал, «проведенческую концепцию» нейрона, и предложил вместо нее системную концепцию интегративной деятельности нейрона.
Вне зависимости от конкретных, усложняющихся с развитием науки представлений о функционировании нейрона, в традиционном рассмотрении центральной оставалась идея об электрической суммации потенциалов на мембране нейрона. В соответствии с ней предполагалось, что возбуждающие и тормозные постсинаптические потенциалы, возникающие за счет изменения ионных градиентов на мембране постсинаптического («получающего») нейрона под действием пресинаптической импульсации, суммируясь, действуют на генераторный пункт нейрона, продуцирующий распространяющиеся потенциалы действия — импульсы.
П. К. Анохин назвал парадоксальным перенос на нейрон представлений, возникших при изучении функционирования нервного волокна и заставляющих рассматривать проведение возбуждения как главную деятельность нервной клетки. Если задача состоит лишь в том, чтобы передать возбуждение от одного нейрона к другому, то не ясно, для чего между входным и выходным импульсами «вставлены» сложные промежуточные этапы: выделение медиатора, его воздействие на субсинаптическую мембрану и химические превращения в ней. «Неужели для того, чтобы, начав с электрического потенциала терминали, сформировать в конце концов тот же спайковый потенциал, весьма сходный по своим физическим параметрам с потенциалом, пришедшим по аксонной терминали?» [Анохин, 1975, с. 368].
14.3.2. Парадигма активности: нейрон, как и индивид,
достигает «результата», получая необходимые
Дата добавления: 2015-06-10; просмотров: 923;