Происхождение эукариот

Появление эукариот – важнейшее событие. Изменило структуру биосферы и открыло принципиально новые возможности для прогрессивной эволюции. Эукариотическая клетка является результатом долгой эволюции мира прокариот, мира, в котором разнообразные микробы приспосабливались друг к другу и искали способы эффективной кооперации.
набросок хронологии (повторение)

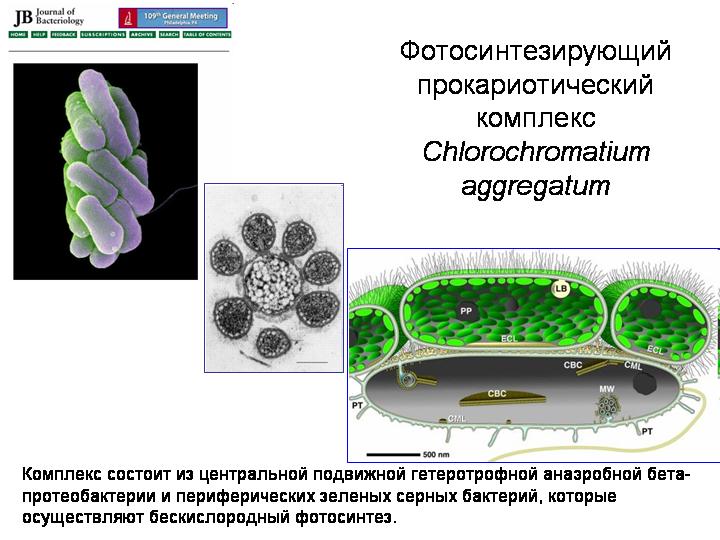
Фотосинтезирующий прокариотический комплекс Chlorochromatium aggregatum.
Эукариоты возникли в результате симбиоза нескольких видов прокариот. Прокариоты вообще весьма склонны к симбиозу (см. главу 3 в книге «Рождение сложности»). Вот интересная симбиотическая система, известная под названием Chlorochromatium aggregatum. Живет в глубоких озерах, где есть на глубине бескислородные условия. Центральный компонент – подвижная гетеротрофная бета-протеобактерия. Вокруг нее стопками располагаются от 10 до 60 фотосинтезирующих зеленых серных бактерий. Все компоненты соединены выростами наружной мембраны центральной бактерии. Смысл содружества в том, что подвижная бета-протеобактерия перетаскивает всю компанию в места, благоприятные для жизни привередливых серных бактерий, а серные бактерии занимаются фотосинтезом и обеспечивают пищей и себя, и бета-протеобактерию. Может быть, какие-то древние микробные ассоциации примерно такого типа и были предками эукариот.
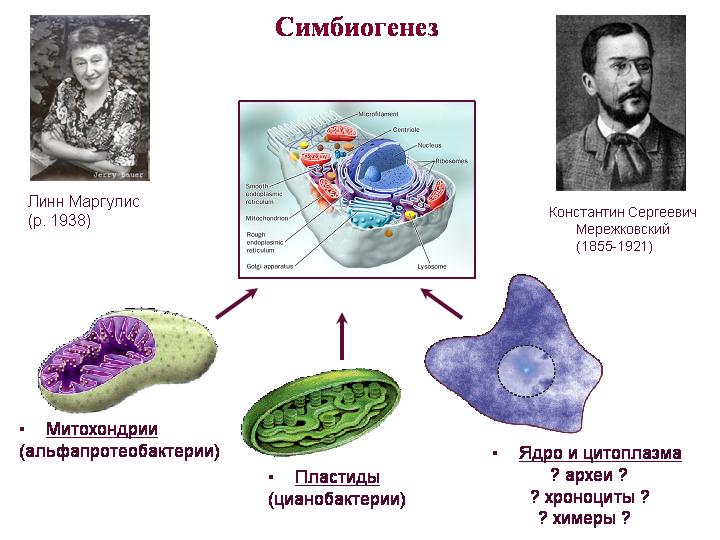
Теория симбиогенеза. Мережковский, Маргулис. Митохондрии – потомки альфа-протеобактерий, пластиды – потомки цианобактерий. Труднее понять, кто был предком всего остального, то есть цитоплазмы и ядра. Ядро и цитоплазма эукариот сочетает в себе признаки архей и бактерий, а также имеет множество уникальных особенностей.
О митохондриях. Возможно, именно приобретение митохондрий (а не ядра) было ключевым моментом в становлении эукариот. Большинство генов предков митохондрий было перенесено в ядро, где они попали под контроль ядерных регуляторных систем. Эти ядерные гены митохондриального происхождения кодируют не только белки митохондрий, но и многие белки, работающие в цитоплазме. Это говорит о том, что митохондриальный симбионт сыграл более важную роль в формировании эукариотической клетки, чем предполагалось.
Совместное существование в одной клетке двух разных геномов требовало развития эффективной системы их регуляции. А для того, чтобы эффективно управлять работой большого генома, необходимо изолировать геном от цитоплазмы, в которой протекает обмен веществ и идут тысячи химических реакций. Ядерная оболочка как раз и отделяет геном от бурных химических процессов цитоплазмы. Приобретение симбионтов (митохондрий) могло стать важным стимулом в развитии ядра и генно-регуляторных систем.
То же относится и к половому размножению. Без полового размножения можно жить до тех пор, пока геном у вас достаточно маленький. Организмы с большим геномом, но лишенные полового размножения, обречены на быстрое вымирание, за редчайшими исключениями.

Альфапротеобактерии – к этой группе относились предки митохондрий.
Родоспириллум – удивительный микроорганизм, который может жить и за счет фотосинтеза, в том числе и в анаэробных условиях, и как аэробный гетеротроф, и даже как аэробный хемоавтотроф. Он может, например, расти за счет окисления угарного газа СО, не используя никаких других источников энергии. Вдобавок ко всему этому, он умеет еще и фиксировать атмосферный азот. То есть это в высшей степени универсальный организм.
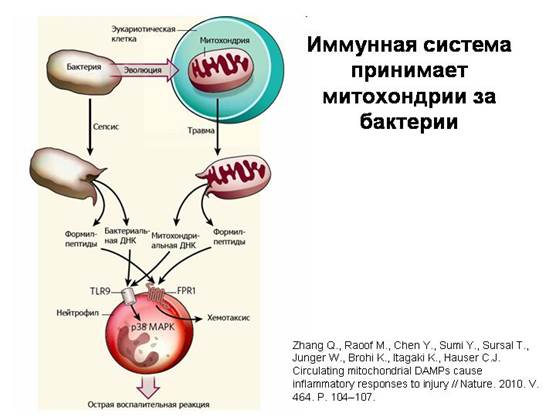
Иммунная система принимает митохондрии за бактерии. Когда при травме в кровь попадают разрушенные митохондрии, из них высвобождаются характерные молекулы, которые встречаются только у бактерий и у митохондрий (кольцевая ДНК бактериального типа и белки, несущие на одном из своих концов особую модифицированную аминокислоту формилметионин). Это связано с тем, что аппарат синтеза белка в митохондриях остался таким же, как у бактерий. Клетки иммунной системы – нейтрофилы – реагируют на эти митохондриальные вещества точно так же, как на бактериальные, и при помощи тех же рецепторов. Это ярчайшее подтверждение бактериальной природы митохондрий.

Главная функция митохондрий – кислородное дыхание. Скорее всего, стимулом для объединения анаэробного предка ядра и цитоплазмы с «протомитохондрией» была необходимость защититься от токсического действия кислорода.
Откуда взялись у бактерий, и в том числе у альфапротеобактерий, молекулярные системы, необходимые для кислородного дыхания? Похоже, в их основу были положены молекулярные системы фотосинтеза. Электронно-транспортная цепь, сформировавшаяся у бактерий как часть фотосинтетического аппарата, была адаптирована для кислородного дыхания. У некоторых бактерий до сих пор участки электронно-транспортных цепей используются одновременно и в фотосинтезе, и в дыхании. Скорее всего предками митохондрий были аэробные гетеротрофные альфа-протеобактерии, которые, в свою очередь, произошли от фотосинтезирующих альфа-протеобактерий, таких как родоспириллум.

Число общих и уникальных белковых доменов у архей, бактерий и эукариот. Белковый домен – это часть белковой молекулы, имеющая определенную функцию и характерную структуру, то есть последовательность аминокислот. Каждый белок, как правило, содержит один или несколько таких структурно-функциональных блоков, или доменов.
4,5 тысячи белковых доменов, которые есть у эукариот, можно разделить на 4 группы: 1) имеющиеся только у эукариот, 2) общие для всех трех надцарств, 3) общие для эукариот и бактерий, но отсутствующие у архей; 4) общие для эукариот и архей, но отсутствующие у бактерий. Мы рассмотрим две последние группы (они на рисунке выделены цветом), поскольку для этих белков можно с определенной уверенностью говорить об их происхождении: соответственно бактериальном или архейном.
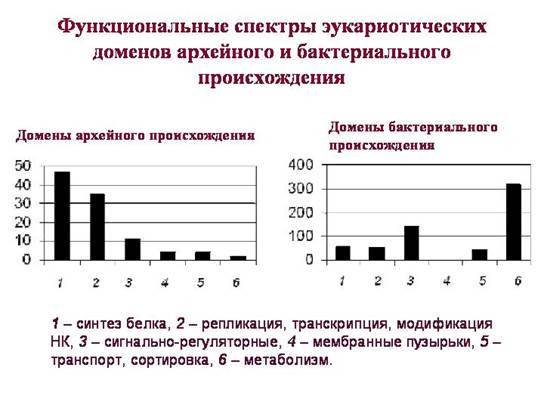
Ключевой момент в том, что эукариотические домены, предположительно унаследованные от бактерий и от архей, имеют существенно разные функции. Домены, унаследованные от архей (их функциональный спектр показан на левом графике), играют ключевую роль в жизни эукариотической клетки. Среди них преобладают домены, связанные с хранением, воспроизведением, организацией и считыванием генетической информации. Большинство "архейных" доменов относится к тем функциональным группам, в пределах которых горизонтальный обмен генами у прокариот происходит реже всего. Видимо, эукариоты получили этот комплекс путем прямого (вертикального) наследования от архей.
Среди доменов бактериального происхождения тоже есть белки, связанные с информационными процессами, но их мало. Большинство из них работает только в митохондриях или пластидах. Эукариотические рибосомы цитоплазмы имеют архейное происхождение, рибосомы митохондрий и пластид имеют бактериальное происхождение.
Среди бактериальных доменов эукариот значительно выше доля сигнально-регуляторных белков. От бактерий эукариоты унаследовали многие белки, ответственные за механизмы реагирования клетки на факторы внешней среды. А также – многие белки, связанные с обменом веществ (подробнее см. в главе 3 «Рождения сложности»).
Эукариоты имеют:
· Архейную «сердцевину» (механизмы работы с генетической информацией и синтеза белка)
· Бактериальную «периферию» (обмен веществ и сигнально-регуляторные системы)
· Простейший сценарий: АРХЕЯ проглотила БАКТЕРИЙ (предков митохондрий и пластид) и все свои бактериальные признаки приобрела от них.
· Этот сценарий слишком прост, потому что у эукариот много бактериальных белков, которые не могли быть заимствованы у предков митохондрий или пластид.
У эукариот много «бактериальных» доменов, не характерных ни для цианобактерий (предков пластид), ни для альфапротеобактерий (предков митохондрий). Они были получены от каких-то других бактерий.


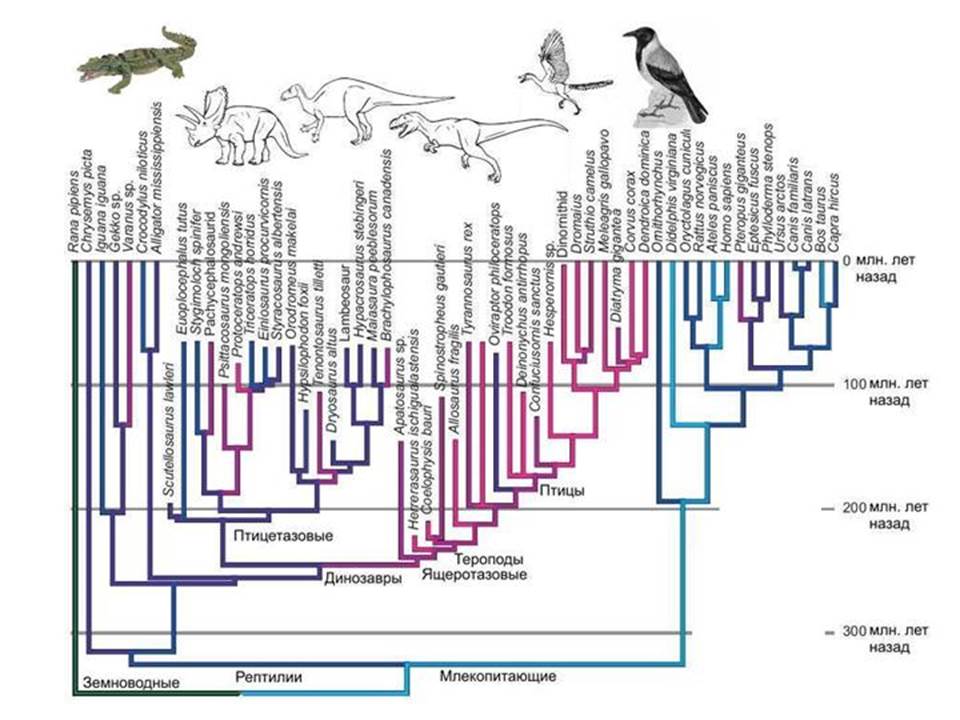
Птицы и динозавры. Реконструировать прото-эукариот трудно. Ясно, что та группа древних прокариот, которая дала начало ядру и цитоплазме, обладала рядом уникальных особенностей, которых нет у прокариот, доживших до наших дней. И когда мы пытаемся реконструировать облик этого предка, мы сталкиваемся с тем, что простор для гипотез оказывается слишком большим.
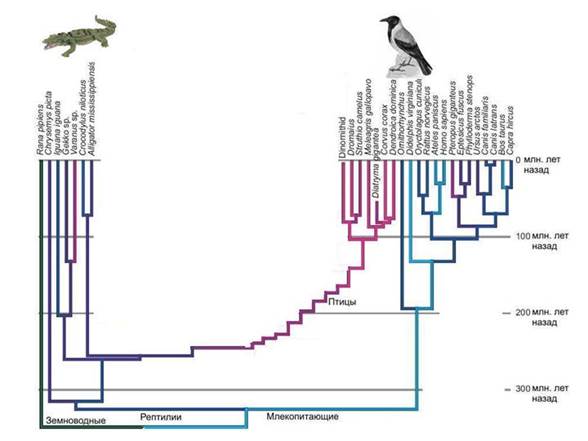
Аналогия. Известно, что птицы произошли от динозавров, причем не от каких-то неизвестных динозавров, а от вполне определенной группы – манирапторых динозавров, которые относятся к тероподам, а тероподы в свою очередь – это одна из групп ящеротазовых динозавров. Найдено много переходных форм между нелетающими динозаврами и птицами.
Но что бы мы могли сказать о предках птиц, если бы ископаемой летописи не было? В лучшем случае мы бы выяснили, что ближайшей родней птиц являются крокодилы. Но смогли бы мы воссоздать облик прямых предков птиц, то есть динозавров? Вряд ли. Но именно в таком положении мы и находимся, когда пытаемся восстановить облик предка ядра и цитоплазмы. Ясно, что это была группа неких прокариотических динозавров, группа вымершая и не оставившая, в отличие от настоящих динозавров, внятных следов в геологической летописи. Современные археи по отношению к эукариотам – это как современные крокодилы по отношению к птицам. Попробуйте восстановить строение динозавров, зная только птиц и крокодилов.
Аргумент в пользу того, что в докембрии жило много всяких микробов, не похожих на нынешние. Протерозойские строматолиты были намного сложнее и разнообразнее современных. Строматолиты – продукт жизнедеятельности микробных сообществ. Не значит ли это, что и протерозойские микробы были разнообразнее современных, и что многие группы протерозойских микробов просто не дожили до наших дней?

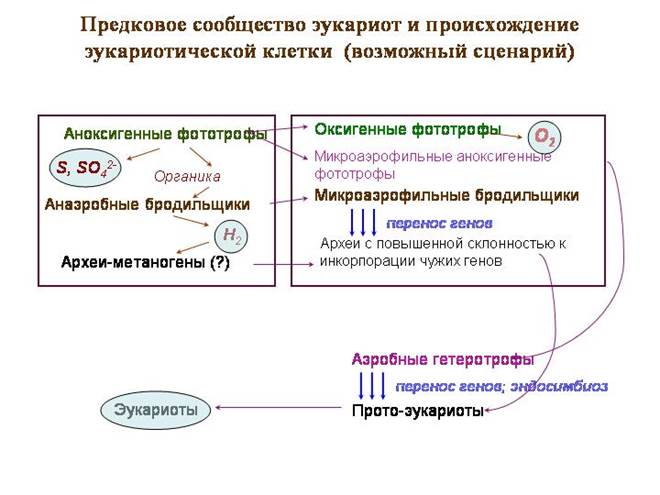
Предковое сообщество эукариот и происхождение эукариотической клетки (возможный сценарий)
Гипотетическое «предковое сообщество» - типичный бактериальный мат, только в его верхнем жили предки цианобактерий, еще не перешедшие к оксигенному фотосинтезу. Они занимались аноксигенным фотосинтезом.Донором электронов служила не вода, а сероводород. В качестве побочного продукта выделялись сера и сульфаты.
Во втором слое обитали пурпурные фотосинтезирующие бактерии, в том числе – альфапротеобактерии, предки митохондрий. Пурпурные бактерии используют длинноволновой свет (красный и инфракрасный). Эти волны обладают лучшей проникающей способностью. Пурпурные бактерии и сейчас часто живут под слоем цианобактерий. Пурпурные альфапротеобактерии тоже используют в качестве донора электрона сероводород.
В третьем слое были бактерии-бродильщики, перерабатывающие органику; некоторые из них в качестве отходов выделяли водород. Это создавало базу для сульфатредуцирующих бактерий. Там могли быть и метаногенные археи. Среди обитавших здесь архей были и предки ядра и цитоплазмы.
Начало кризисным событиям положил переход цианобактерий к кислородному фотосинтезу. В качестве донора электрона цианобактерии начали использовать вместо сероводорода обычную воду. Это открывало большие возможности, но имело и негативные последствия. Вместо серы и сульфатов при фотосинтезе стал выделяться кислород – вещество крайне токсичное для всех древних обитателей земли.
Первыми с этим ядом столкнулись его производители – цианобактерии. Они же, вероятно, первыми стали вырабатывать средства защиты от него. Электронно-транспортные цепи, служившие для фотосинтеза, были модифицированы и начали служить для аэробного дыхания. Изначальная цель, видимо, заключалась не в получении энергии, а только в нейтрализации кислорода.
Вскоре и обитателям второго слоя сообщества – пурпурным бактериям – пришлось вырабатывать аналогичные системы защиты. Так же, как и цианобактерии, они сформировали системы аэробного дыхания на основе фотосинтетических систем. Именно у пурпурных альфапротеобактерий развилась наиболее совершенная дыхательная цепь, которая ныне функционирует в митохондриях эукариот.
В третьем слое сообщества появление свободного кислорода должно было вызвать кризис. Метаногены и многие сульфатредукторы утилизируют молекулярный водород при помощи ферментов-гидрогеназ. Такие микробы не могут жить в аэробных условиях, потому что кислород ингибирует гидрогеназы. Многие бактерии, выделяющие водород, в свою очередь, не растут в среде, где нет микроорганизмов, его утилизирующих. Из бродильщиков в составе сообщества, по-видимому, остались формы, выделяющие в качестве конечных продуктов низкоорганические соединения (пируват, лактат, ацетат и т.п.). Эти бродильщики выработали свои средства защиты от кислорода, менее эвффективные. К числу выживших относились и археи – предки ядра и цитоплазмы.
Может быть, в этот кризисный момент и произошло ключевое событие – ослабление генетической изоляции у предков эукариот и начало активного заимствования чужих генов. Прото-эукариоты инкорпорировали гены разных бродильщиков до тех пор, пока не стали сами микроаэрофильными бродильщиками, сбраживающими углеводы до пирувата и молочной кислоты.
Обитатели третьего слоя – предки эукариот – теперь непосредственно контактировали с новыми обитателями второго слоя – аэробными альфапротеобактериями, которые научились использовать кислород для получения энергии. Метаболизм прото-эукариот и альфапротеобактерий стал взаимодополнительным, что создавало предпосылки для симбиоза. Да и само расположение альфапротеобактерий в сообществе (между верхним, выделяющим кислород, и нижним слоем) предопределяло их роль как «защитников» предков эукариот от избытков кислорода.
Вероятно, прото-эукариоты заглатывали и приобретали в качестве эндосимбионтов многих разных бактерий. Экспериментирование такого рода и сейчас продолжается у одноклеточных эукариот, обладающих огромным разнообразием внутриклеточных симбионтов. Из этих экспериментов союз с аэробными альфапротеобактериями оказался наиболее удачным.
Происхождение эукариот: заключение
· От архей эукариоты унаследовали многие компоненты информационных систем нуклеоцитоплазмы.
· Бактериальные эндосимбионты внесли большой вклад в формирование метаболических и сигнально-регуляторных систем не только в органеллах, но и в цитоплазме.
· Еще до приобретения эндосимбионтов прото-эукариоты заимствовали у разных бактерий ряд генных комплексов с метаболическими и сигнально-регуляторными функциями.
· По-видимому, в эволюции прото-эукариот был период ослабления изоляционных барьеров. В это время происходила активная инкопрорация чужих генов.
· В роли «спускового крючка» цепочки событий, приведших к появлению эукариот, выступил кризис микробных сообществ, вызванный переходом цианобактерий к кислородному фотосинтезу.









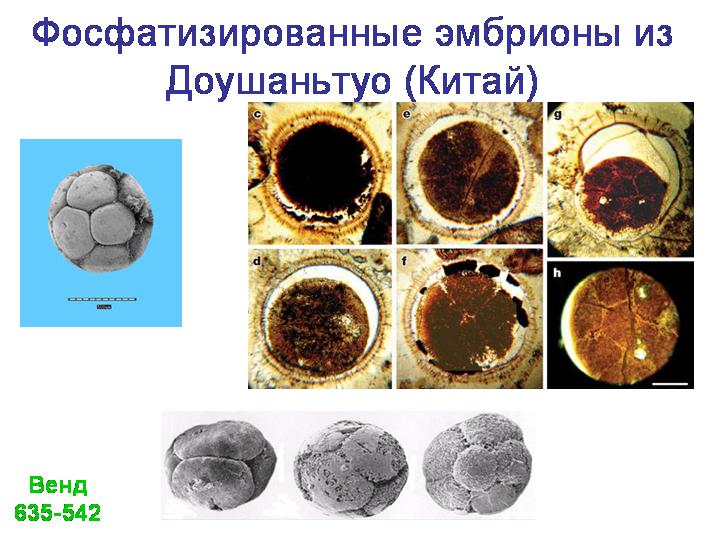
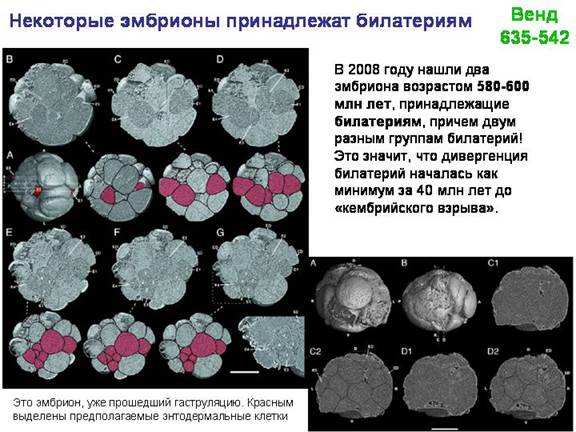


















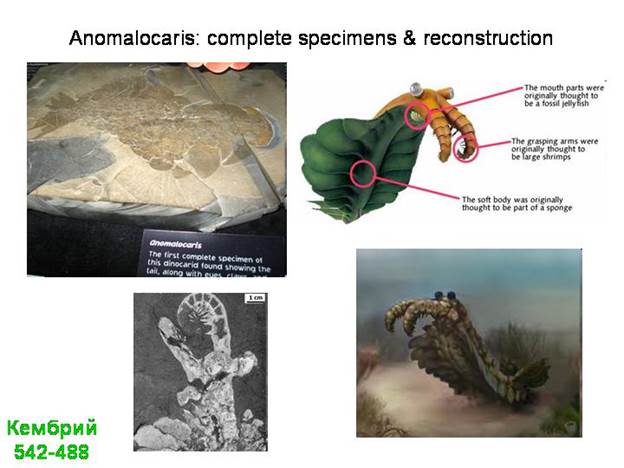


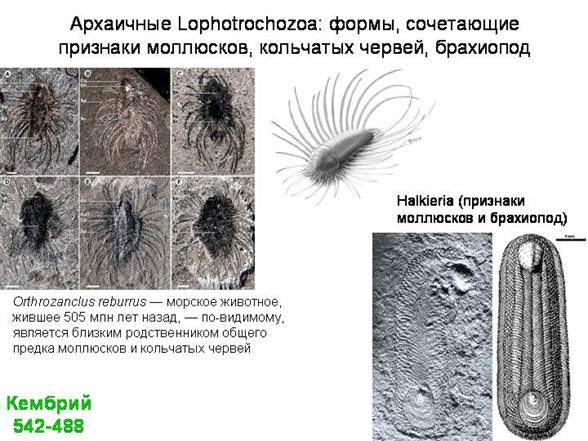
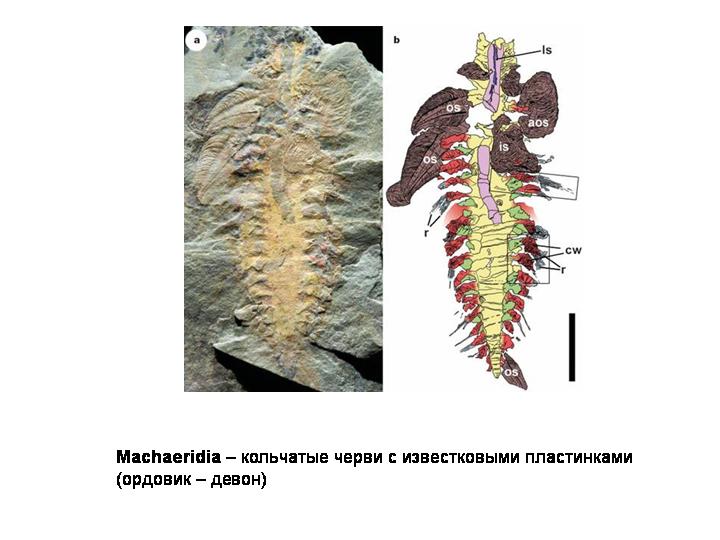





















В качестве одной из лекций - экскурсия в Палеонтологический музей (обязательно!).
Часть 8. Палеоантропология (лекции 26-28)
Дата добавления: 2015-06-10; просмотров: 1743;