Горизонтальный перенос генов
Вертикальная передача генов – от родителей к потомкам. Горизонтальная – от одного организма другому, не являющемуся его потомком.

Прочтение геномов множества прокариот и эукариот показало, что традиционные представления об эволюции, базирующиеся на идее дивергенции (расхождения видов), недостаточны для понимания генеалогии видов. На этом слайде показана знаменитая дарвиновская схема – единственный рисунок, которым Дарвин снабдил главный свой труд «Происхождение видов». Ветви расходятся, чтобы больше никогда не соединиться.
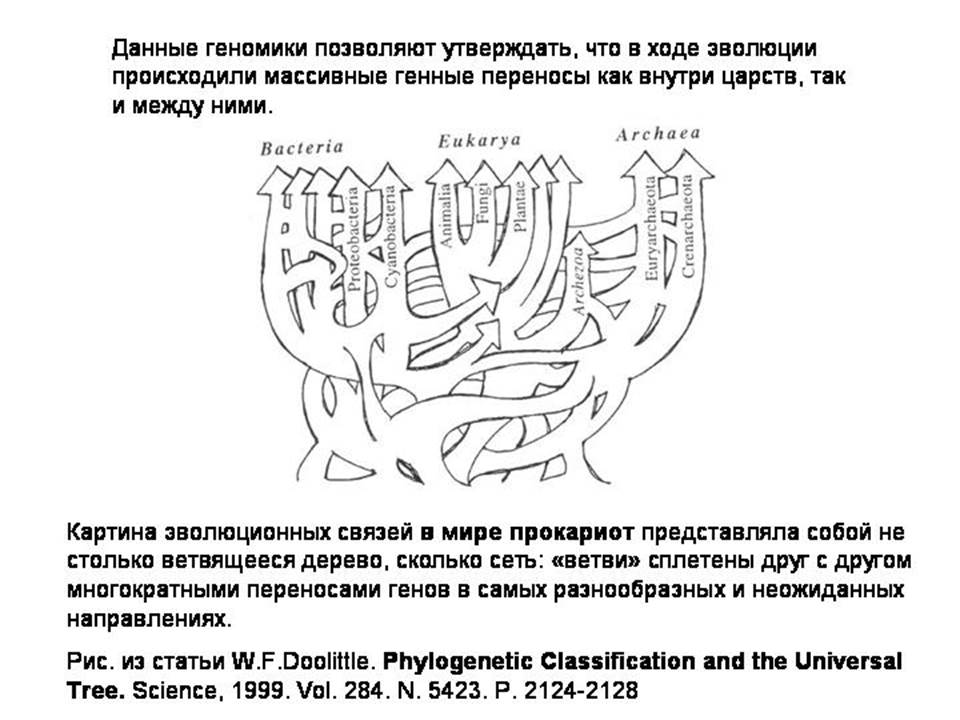
Данные геномики (сравнения геномов) позволяют утверждать, что в ходе эволюции многократно происходили генные переносы между разными группами организмов. По-видимому, на самых ранних этапах эволюции существовало некое общее генное "коммунальное хозяйство". Картина эволюционных связей в мире предковых (древних) прокариот представляла собой не столько дерево, сколько сеть (дерево с горизонтальными перемычками между ветвями).
По мере усложнения организмов и развития механизмов полового размножения горизонтальный перенос становился более редким явлением.

Горизонтальный перенос генов можно выявить по ряду показателей. Во-первых, по нуклеотидному составу ДНК. Отличие в нуклеотидном составе отдельного сегмента от остальной части генома является указанием на присутствие "чужих" генов. Во-вторых, по частоте встречаемости в гене определенных кодонов. Третий важный критерий – существенное отличие в положении анализируемого гена на филогенетическом (эволюционном) дереве от большинства других генов. О "чужеродном" происхождении гена может говорить высокая степень его сходства с гомологичным (т.е. соответствующим) геном из отдаленного таксона при отсутствии подобного гена у близких "родственников".
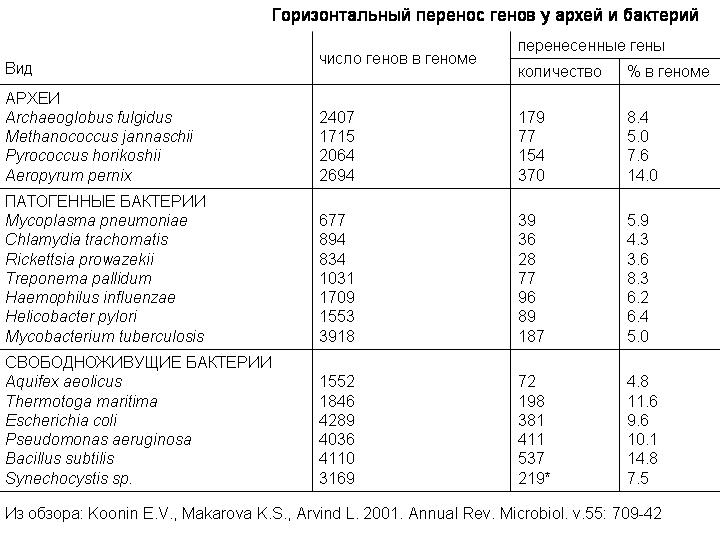
Это таблица из обзора 2001 года, которая дает представление о масштабах горизонтального переноса у прокариот. Примерно от 4 до 15 процентов генов были получены разными видами микробов путем горизонтального переноса. Речь здесь идет прежде всего о недавних событиях, потому что древние акты горизонтального переноса выявить сложнее. По последним данным число перенесенных генов может быть существенно больше. Вот здесь в таблице написано, что у E.coli 381 перенесенный ген, или 9,6%. А по более новым данным у нее 755 перенесенных генов, то есть 18%, вдвое больше. Эти 755 генов были приобретены в течение последних 200 млн лет. Примерно по 4 гена за миллион лет.
Некоторые общие закономерности горизонтального переноса у прокариот:
· Доля латерально полученных генов варьирует у разных видов и может достигать 10-15%. По последним данным, может быть и больше.
· Наибольшее количество переносов характерно для свободноживущих бактерий с широкими экологическими ареалами.
· Наименьшее число переносов обнаружено у патогенных бактерий, живущих в узких эконишах.
· Реже всего в горизонтальные переносы вовлечены гены информационных систем (транскрипции, трансляции, репликации), составляющие базовый геном. Продукты этих генов входят в сложные белковые комплексы, где "чужие" белки не встраиваются или не функционируют.
· Чаще всего в горизонтальном переносе участвуют гены, связанные с метаболизмом, транспортными путями и передачей сигналов.
· В составе приобретенных сегментов ДНК часто обнаруживаются профаги, плазмиды, гены белков, участвующих в процессах рекомбинации.
Горизонтальная передача генов реализуется через различные каналы генетической коммуникации.
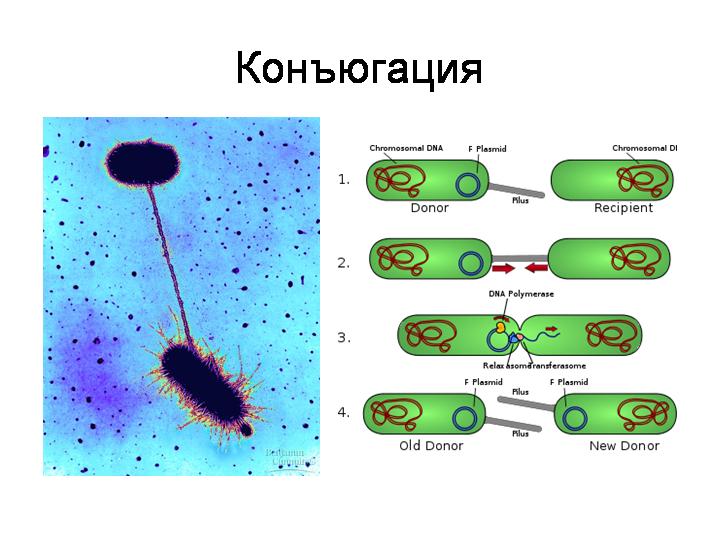
· Коньюгация (одна бактерия при помощи специальной трубки впрыскивает часть своей ДНК в другую бактерию)
· Трансформация (бактерия просто заглатывает ДНК из внешней среды)
· Трансдукция (перенос фрагментов ДНК вместе с вирусами)
· Перенос генов может происходить в симбиотических и паразитарных системах, где есть физический контакт клеток с разными геномами.
· «Случайное» включение чужих генов в ходе репарации разрывов ДНК, – этот механизм, по-видимому, важен для некоторых многоклеточных эукариот.
· И, наконец, половой процесс, который обычно не рассматривается как вариант горизонтального генетического обмена, хотя суть явления в принципе та же самая: это объединение в одном геноме генов разных организмов. В обоих случаях речь идет, если можно так выразиться, о межорганизменной генетической рекомбинации. Т.е. комбинирование генов разных организмов, обмен участками генома между разными организмами.
Судя по всему, появление полового процесса было закономерным результатом эволюции механизмов межорганизменной рекомбинации.
Половой процесс отличается от более примитивных механизмов двумя основными чертами:
1) большей избирательностью (то есть смешение генов происходит в основном между близкородственными организмами). Однако и у прокариот, конечно, есть такая избирательность: близкородственные микробы обмениваются генами намного чаще, чем филогенетически удаленные. У эукариот избирательность стала настолько строгой, что это привело к появлению относительно замкнутых группировок – репродуктивно изолированных, эндогамных группировок – которые мы называем биологическими видами.
У прокариот таких строго эндогамных группировок нет. Поэтому некоторые теоретики предлагают даже считать всех прокариот одним биологическим видом.
2) Второе отличие – при половом процессе объединяются и рекомбинируют не фрагменты геномов, а целые геномы. Эти отличия (между горизонтальным генетическим обменом и половым процессом) конечно очень важные, но в определенном смысле они не столько качественные, сколько количественные.

Современная генная инженерия, по сути дела, базируется на принципах горизонтального переноса генов, хотя еще недавно не было четкого понимания того, что такого рода генная инженерия широко распространена в природе.
Действительно, например, бактерии, способные к конъюгации, являются самыми настоящими природными генными инженерами. Причем они могут вводить свою ДНК не только в клетки других прокариот, но и в эукариотические клетки, как показывает пример агробактерии. Агробактерия вводит часть своей ДНК в растительную клетку, что приводит к развитию опухоли, в которой бактерия чувствует себя комфортно. Характерно, что агробактерия, как и другие родственные ей микробы, использует для введения своей ДНК или белков в клетки эукариот модифицированный аппарат конъюгации. Конъюгацию называют аналогом полового процесса у прокариот. И получается, что агробактерия совершает с растительными клетками нечто похожее на половой процесс.
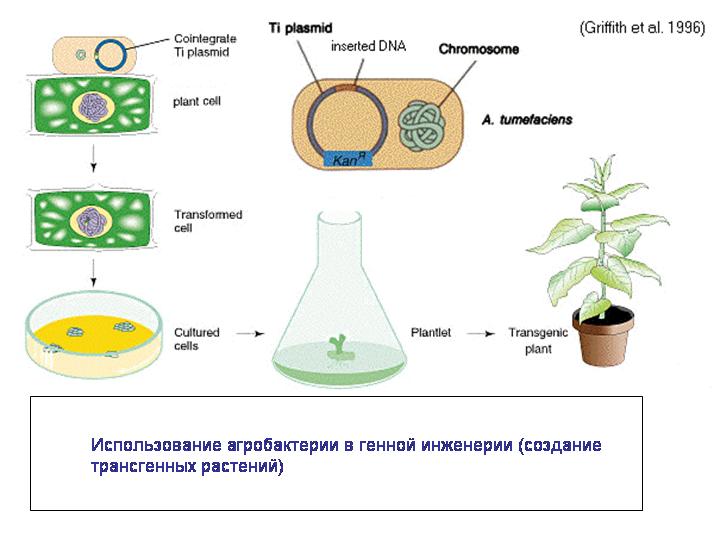
Агробактерия сейчас широко используется для создания трансгенных растений. Так что генная инженерия – это вовсе не человеческое изобретение, мы научились этому у природы и пользуемся природными технологиями.
Это были общие, начальные сведения о горизонтальном переносе генов. А теперь – несколько недавно установленных фактов.
Начнем с прокариот.
 .
.
Сотрудники институт Крейга Вентера показали, что у прокариот в принципе возможен обмен целыми геномами, что может приводить к превращению одного вида бактерий в другой.
Этот институт работает над созданием искусственных микробов. Например, планируется создание микробов, которые будут производить дешевое топливо. Работы ведутся с бактериями рода Mycoplasma. Одна из важных задач проекта – это научиться вставлять целый геном в живую бактерию, полностью заменив при этом ее собственный геном.
В 2007 г Вентер и его коллеги сообщили о первой успешной пересадке целого генома от одного вида бактерий другому. Сделано было следующее. Выделили геном из бактерии Mycoplasma mycoides, которая вызывает пневмонию у коров. Геном был очищен от всех примесей и добавлен в культуру бактерий Mycoplasma capricolum, возбудителей козьего полиартрита. Эти два вида микоплазм довольно близкие, разошлись они несколько десятков миллионов лет назад, может быть, тогда же, когда разошлись их хозяева – коровы и козы. На слайде красным кружком показано положение этих видов на эволюционном древе класса Mollicutes.
Вскоре среди клеток Mycoplasma capricolum появились бактерии с признаками Mycoplasma mycoides. Детальный анализ показал, что эти бактерии по всем признакам – самые настоящие M. mycoides. Один вид бактерий превратился в другой.
По-видимому, бактерии Mycoplasma capricolum «проглатывали» чужую молекулу ДНК, т.е. имела место трансформация.
Сначала в них содержались оба генома вместе. Когда такая клетка делилась, одна из дочерних клеток получала геном одного вида, другая – другого.
Этот эксперимент показал, что у прокариот в принципе возможны и такие полногеномные акты горизонтального переноса. То есть при трансформации у прокариот в принципе нет жестких ограничений на размер фрагментов ДНК, получаемых из внешней среды. Могут передаваться не только маленькие фрагменты, но и большие, и даже целые геномы. Происходит ли такое в природе – не известно (вряд ли).
Пример, показывающий, для чего может быть нужен прокариотам горизонтальный обмен генами.
У некоторых морских планктонных бактерий известны белки – протеородопсины, позволяющие частично утилизировать энергию солнечного света. Это гораздо более простая система, чем настоящий фотосинтез, хотя и менее эффективная. Но для ее работы достаточно всего одного или двух генов, поэтому эта система легко может передаваться путем горизонтального переноса. Оказалось, что так оно и происходит. Протеородопсины обнаружены не только у разных групп бактерий в фотической зоне океана, но и у многих архей, тоже живущих в фотической зоне. Причем эти гены у бактерий и архей очень похожи друг на друга, поэтому ясно, что они передаются от одного вида другому. Получается, что эти гены являются неким общим достоянием, коллективной собственностью сообщества прокариот, живущих в фотической зоне океана.
Каков в целом масштаб явления?
Ранее мы говорили о генах, которые были перенесены сравнительно недавно, и поэтому факт переноса нетрудно заметить, следы еще не затерлись.

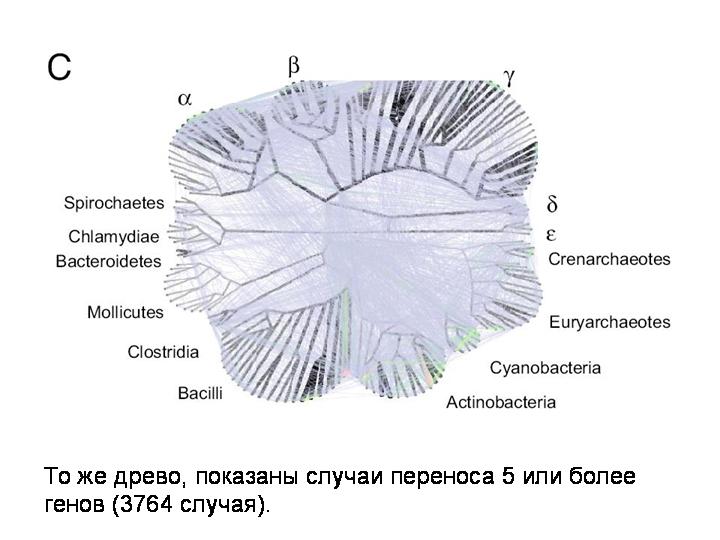
Очень интересная статья об этом вышла в июле нынешнего года. К 180 прокариотическим геномам были применены новые математические методы анализа, разработанные для изучения сетевых структур. Примененная методика позволила выявить не только недавние события горизонтального переноса, то есть связи между концевыми ветвями эволюционного древа, но и древние события, которые связывают между собой все узлы эволюционного древа прокариот.
И вот что получилось. На этом слайде показано базовое филогенетическое древо, основанное на генах рРНК.
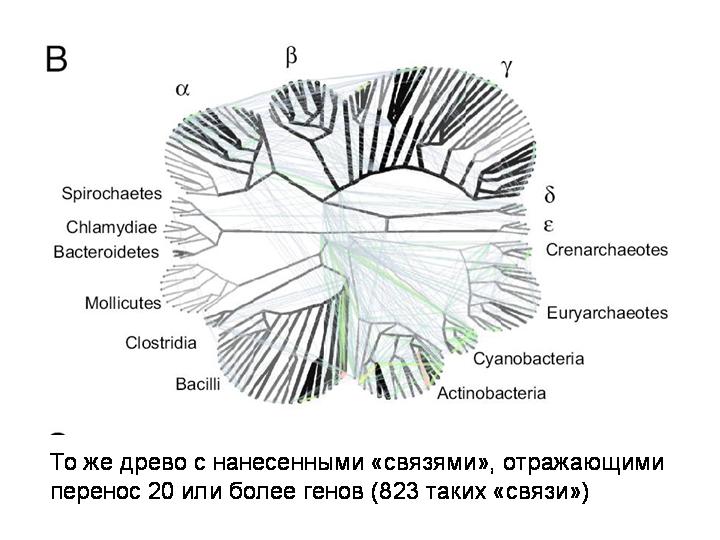
То же древо с нанесенными «связями», отражающими перенос 20 или более генов. Всего выявлено 823 случая, когда из одной точки эволюционного дерева прокариот в другую точку было горизонтально перенесено 20 или более генов.
 .
.

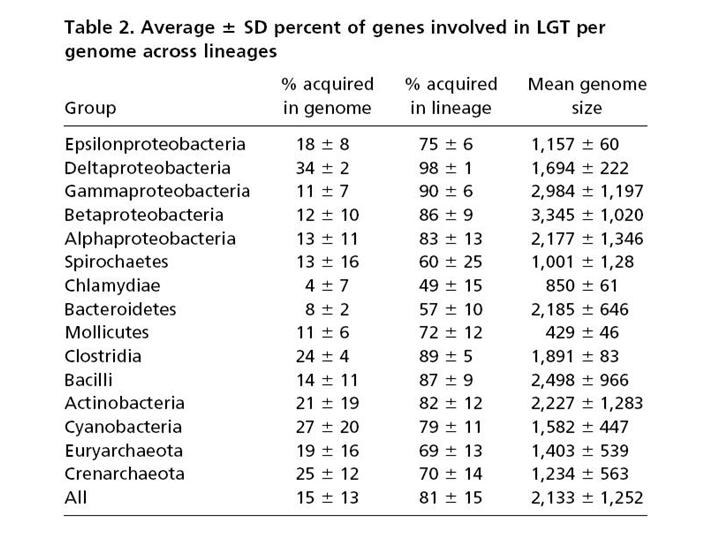
Вот цифры. В первой колоке – процент генов, приобретенных путем горизонтального переноса в среднем на геном, если учитывать только те гены, которые были приобретены недавно, то есть после последней дивергенции (последнего разветвления дерева). Мы видим, что цифры примерно такие же или немного больше, чем в той таблице, которую я показывал до этого.
Интереснее вторая колонка цифр. Она показывает процент генов, попавших в данный геном путем горизонтального переноса за всю историю данной эволюционной линии. И здесь мы уже видим цифры, доходящие до 98%.
Не удивительно, что авторы делают вывод о том, что представление эволюции прокариот в виде ветвящегося древа уже нельзя считать адекватным. На самом деле это не дерево, а сеть. Горизонтальный перенос происходит не очень часто, поэтому мы и видим сравнительно небольшой процент недавно приобретенных генов. Однако с течением эволюционного времени кумулятивный, суммарный эффект горизонтального переноса накапливается как снежный ком. На этом слайде показана трехмерная реконструкция эволюции прокариот с учетом горизонтального переноса.
Вот такие поистине глобальные масштабы имеет горизонтальный перенос в мире прокариот.
Ну а что же с эукариотами?
Я не буду (пока) останавливаться на том, что эукариотический ядерный геном является химерным (смешанным) с самого начала. Мы видим в нем смесь генов архейного и бактериального происхождения, которые объединились на ранних этапах становления эукариотической клетки. Потому что эукариотическая клетка развилась в результате слияния нескольких разных прокариот, предком эукариотической клетки было сообщество прокариот. Об этом мы поговорим позже. Но надо понимать, что смешение генов нескольких разных прокариот в едином эукариотическом геноме – это событие, конечно, тоже, следует классифицировать как акт горизонтального переноса, смешивания генов разных организмов в одном геноме.
Сейчас мы поговорим о тех событиях, котрые происходили уже после становления эукариотической клетки. У эукариот нет специальных механизмов для передачи своих генов неродственным организмам. Вроде бы нет и прямых аналогов трансформации. Однако эукариоты, особенно одноклеточные, могут тем не менее заимствовать гены у бактерий. И чем больше прочтенных эукариотических геномов, тем яснее это становится.
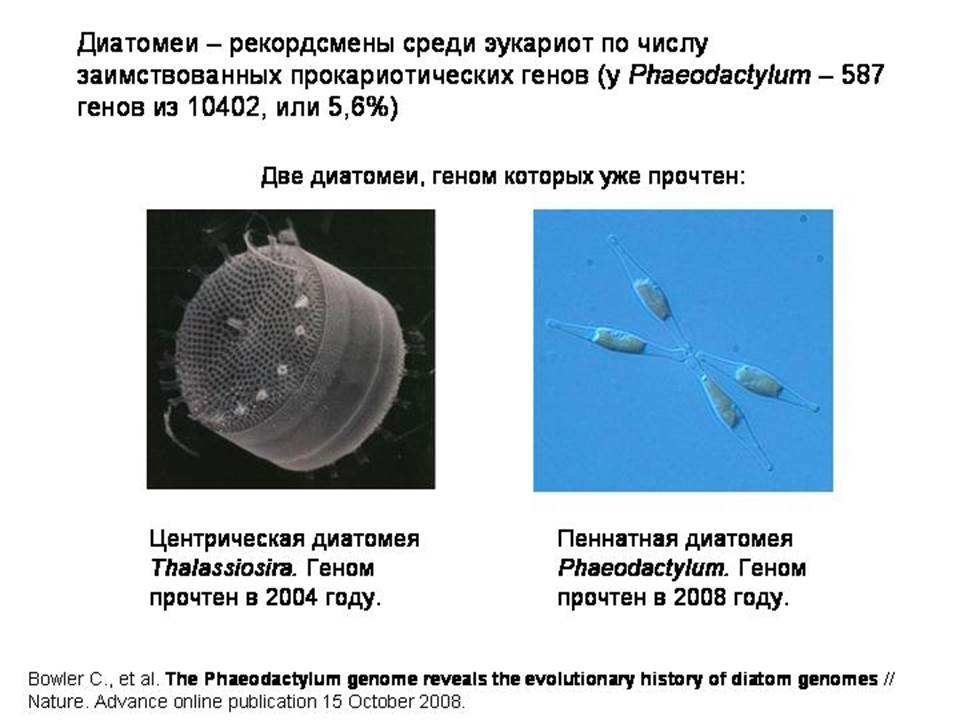
Рекордсменом на сегодняшний день является одноклеточная диатомовая водоросль Phaeodactylum, геном которой был прочтен в 2008 году. В геномах диатомей обнаружено много генов, которые были заимствованы у различных прокариот: цианобактерий, протеобактерий, архей и других. В геноме Phaeodactylum обнаружено 587 таких заимствованных генов, что составляет 5,6% от общего числа генов в геноме. Более половины из этих генов (56%) есть также и у центрической диатомеи Thalassiosira. Эти гены, очевидно, были заимствованы диатомеями у прокариот довольно давно — еще до расхождения эволюционных линий центрических и пеннатных диатомей. А диатомеи появились 180 млн лет назад, в юрском периоде – это довольно молодая группа. Остальные 44% прокариотических генов Phaeodactylum, то есть 258 генов, были заимствованы предками Phaeodactylum уже после разделения линий центрических и пеннатных диатомей, то есть в течение последних 90 млн лет. Отсюда можно подсчитать среднюю скорость заимствования: примерно по три гена за миллион лет. Если помните, у кишечной палочки эта скорость примерно такая же.
Какую пользу получают диатомеи от бактериальных генов? Во-первых, это расширяет их биохимические возможности. В частности, бактериальные гены участвуют в построении ажурных кремневых раковинок — главной «визитной карточки» диатомей. Диатомеи позаимствовали у бактерий также многие гены рецепторных и сигнальных белков, при помощи которых бактерии, а теперь и диатомеи, воспринимают сигналы из окружающей среды и реагируют на них. Среди заимствованных у бактерий рецепторов есть даже несколько светочувствительных белков, благодаря которым диатомеи могут реагировать на изменения освещенности.
Вполне возможно, что активный обмен генами между диатомеями и бактериями был одной из главных причин быстрой эволюции диатомей и их эволюционного успеха.
Следует упомянуть еще один важный момент, связанный с геномами диатомей. Диатомеи вместе с бурыми и золотистыми водорослями входят в состав группы Heteroconta. Считается, что гетероконты появились около 1 млрд лет назад в результате симбиоза гетеротрофной эукариотической клетки с одноклеточной красной водорослью. У красных водорослей, как и у зеленых растений, хлоропласты являются первичными, то есть происходят напрямую от симбиотических цианобактерий. Предки гетероконт проглотили одноклеточную красную водоросль и превратили ее в фотосинтезирующего симбионта. Впоследствии от клетки красной водоросли почти ничего не осталось, кроме внешней оболочки и хлоропласта. Поэтому хлоропласты гетероконт являются «вторичными», и они окружены не двумя, а четырьмя мембранами. Чтобы проверить эту теорию, был предпринят поиск в геномах диатомей генов, похожих на гены красных водорослей. Поиск увенчался успехом: выявили более 170 генов, унаследованных предками диатомей от красных водорослей. Таким образом, массовый горизонтальный перенос генов, связанный с симбиогенезом, происходил и после того, как сформировалась эукариотическая клетка.
В целом новые данные говорят о том, что горизонтальный генетический обмен играет в эволюции одноклеточных эукариот более важную роль, чем считалось до сих пор.
Но вот когда мы переходим от одноклеточных эукариот к многоклеточным, то факты горизонтального переноса сразу становятся гораздо более редкими. И все-таки такие факты есть, и их количество быстро растет.
Растения.

Раффлезия — паразитическое растение с самыми крупными в мире цветами, но полностью лишенное листьев, стебля и корней. Привлекает опылителей запахом тухлого мяса. Раффлезия паразитирует на лианах рода Tetrastigma. Она образует в тканях хозяина тяжи, напоминающие грибной мицелий, и все, что осталось у нее от цветкого растения – это цветок. Систематическое положение раффлезии по морфологическим признакам не определялось, и только в 2007 году по молекулярным данным установили, что она относится к семейству молочайных. По всем генам она группируется с молочайными, однако по одному из митохондриальных генов (nad1B-C) раффлезия оказывается ближайшим родственником своего хозяина – лианы Tetrastigma. Очевидно, часть митохондриального генома была заимствована раффлезией у хозяина.

В последние два-три года было описано еще несколько десятков случаев горизонтального переноса митохондриальных генов от одного растения к другому. Почему-то именно растительные митохондрии активно участвуют в горизонтальном переносе. Часто в обмене участвуют паразитические или эпифитные растения, то есть нужен тесный физический контакт. Рекордсменом является очень примитивное цветковое растение Amborella, которая растет в Новой Каледонии. Она заимствовала у других растений целых 24 митохондриальных гена.
Но описаны и единичные случаи обмена ядерными генами. Например, недавно была обнаружена горизонтальная передача транспозона между двумя видами злаков (рис и просо).
Животные.
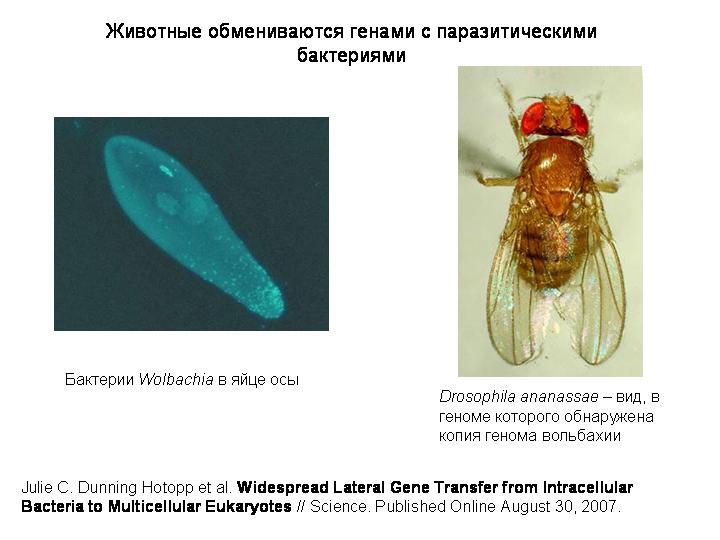
В 2007 году было показано, что гены и даже целые геномы паразитических бактерий иногда могут вставляться в хромосомы животных-хозяев.
Вольбахия — паразитическая бактерия, живущая в клетках многих беспозвоночных. Вольбахию называют микробом-манипулятором, потому что она научилась при помощи регуляторных белков управлять размножением и развитием своим хозяев. Например, она умеет превращать самцов в самок, избирательно убивать зародышей мужского пола, повышать плодовитость зараженных самок и так далее. Вольбахия паразитирует в клетках беспозвоночных уже 100 миллионов лет, и ее предки тоже были внутриклеточными паразитами. При таком долгом и тесном сожительстве было бы даже странно, если бы фрагменты генома вольбахии не попадали бы хотя бы иногда в хозяйский геном. Но до сих пор доказать это не удавалось, и удалось только в 2007 году.
В геномах 4 видов насекомых и 4 видов круглых червей-филярий обнаружены фрагменты генома вольбахии, причем в одном случае - у Drosophila ananassae – геном бактерии оказался вставлен в геном хозяина целиком. Получается, что в ядрах клеток этих мух содержится генетическая информация сразу о двух разных организмах!
Многие гены, заимствованные мухой у бактерии, работают, или по крайней мере транскрибируются. Это значит, что инкорпорация бактериальной ДНК может быть одним из способов приобретения новых генов в эволюции животных.
Теоретически такие бактерии, как вольбахия, паразитирующие в клетках животных, может служить вектором (переносчиком), обеспечивающим передачу генов от одних животных к другим. Ведь в геноме самой вольбахии есть гены, заимствованные у эукариотических хозяев.
.«Молекулярное одомашнивание» можно рассматривать как вариант горизонтального генетического обмена (животные и растения заимствуют полезные гены у вирусов или МГЭ).
Часть 3. Дрейф, отбор, половое размножение (лекции 8-10)
ДЗ. Прочесть «Происхождение видов» Дарвина. Отнестись с уважением, но критически. Написать эссе (1000 – 1500 слов) на тему «Взгляды Дарвина на ......................... с точки зрения современной науки». Вместо многоточия нужно вписать любую интересную и не слишком частную проблему, о которой идет речь в книге. Примеры таких проблем: наследственность, изменчивость, искусственный отбор, естественный отбор, борьба за существование, наследование результатов упражнения органов, половой отбор, происхождение сложных органов, соотношение постепенности и скачкообразности в эволюции, проблема переходных форм, проблема вида в биологии, развитие адаптаций, причины несовершенства организмов, биогеографические следствия эволюции и т.д. и т.п.). Это должен быть аналитический обзор, т.е. пересказ идей Дарвина (по тексту книги, так, чтобы было понятно, что вы ее читали) + критический анализ этих идей с точки зрения того, что нам известно сегодня. Можно опираться на любые заслуживающие доверия источники.
Как правило, мысли Дарвина относятся к одной из следующих категорий:
· Верные, хорошо обоснованные выводы, до сих пор почти или вовсе не устаревшие.
· Гениальные догадки, верные, хотя и основанные на недостаточных (с нашей точки зрения) исходных данных.
· Заблуждения, порожденные отсутствием необходимых фактов (не было генетики, молекулярной биологии и т.д.),
· «Уступки», сделанные (напрасно) под давлением критики.
«Origin of species by means of Natural Selection and all sorts of other things»
Дата добавления: 2015-06-10; просмотров: 2307;