Хлоропласты
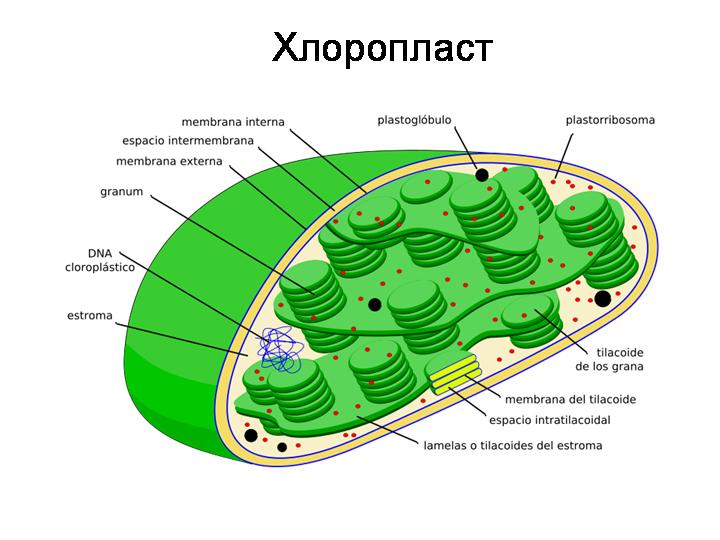
Произошли от симбиотических цианобактерий. Функция – фотосинтез (оксигенный). Имеют собственную кольцевую хромосому, свои рибосомы, самостоятельно размножаются делением, как бактерии. Органы фотосинтеза растений. Кроме цианолбактерий и их потомков – хлоропластов никто не умеет осуществлять кислородный фотосинтез.
Фотосинтез (оксигенный)
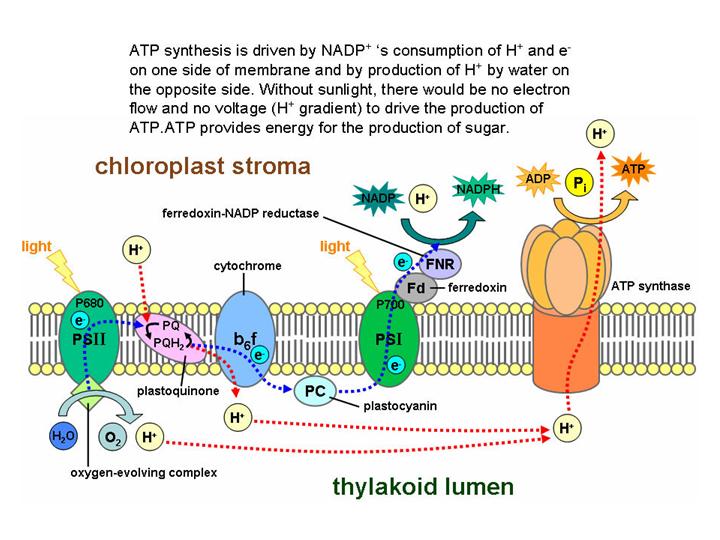

Электронно-транспортная цепь, расположенная на внутренней мембране хлоропласта (= на мембранах тилакоидов). Похожа на Э-Т Ц окислительного фосфорилирования, только электрон идет в обратную сторону. При окислительном фосфорилировании электрон движется в «естественную» сторону, «вниз», постепенно отдавая энергию и в конце концов попадает на молекулу кислорода, которая превращается в воду. При фотосинтезе электрон время от времени (дважды) «подскакивает вверх» за счет энергии солнечного света, а между этими подскоками идет «вниз», отдавая энергию. Эта энергия используется сначала для создания электрохимического градиента, а потом для синтеза АТФ – так же, как при окислительном фосфорилировании.
Электрон изначально берется у воды, и в результате образуется кислород. Отдается электрон в конечном счете веществу NADP (близкое к NAD), которое в результате превращается в «энергетически ценное» вещество NADPH, переносчик протонов и электронов.
Это – световая фаза фотосинтеза. Ее результат – синтез АТФ и NADPH, энергетически ценных молекул, которые затем (в темновой фазе) используются для фиксации CO2.
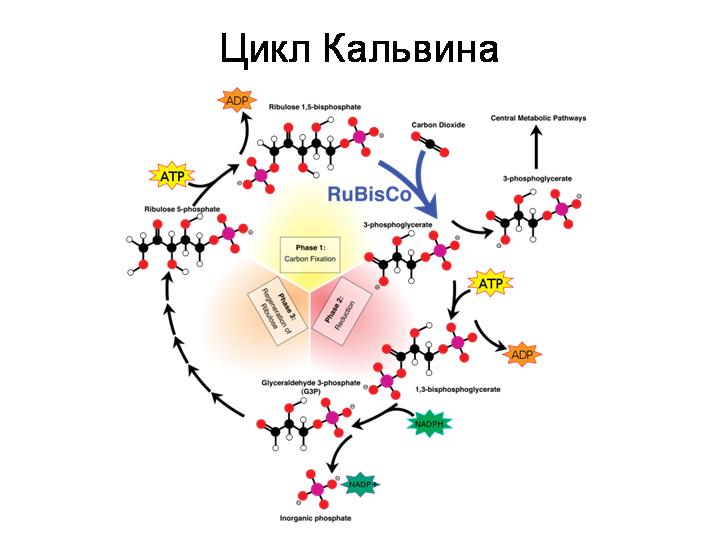
Цикл Кальвина – возможно, самый главный биохимический путь из всех. Это тот способ, при помощи которого живые существа производят органические вещества из неорганического углекислого газа. Присоединить к себе молекулу углекислого газа в состоянии только одно органическое вещество – рибулозодифосфат. Это пятиуглеродный сахар, производное рибозы. Присоединив молекулу CO2, эта молекула из пятиуглеродной превращается в две трехуглеродные молекулы (3-фосфоглицерат). Они потом используются для 1) пополнения запасов рибулозодифосфата, 2) синтеза всех остальных органических веществ, которые нужны клетке. У архей рибулозодифосфат производится из рибонуклеотидов.
Часть 2. Догмы, мобильные элементы и горизонтальный перенос (лекции 5-7)
Задание на следующее занятие: прочесть главу 8 из «Рождения сложности». И приготовить вопросы, если что-то непонятно.
Был задан вопрос «если интроны не нужны, почему они не исчезли, почему отбор их не отсеял»
· Некоторые интроны представляют собой активные МГЭ – они сами размножаются, продуцируют свои копии и встраиваются в новые места хозяйского генома. Т.е. они активно размножаются. Что такое МГЭ и как они устроены, мы скоро разберемся.
· Для большинства интронов эукариот механизм размножения не известен, однако известно, что иногда они все-таки размножаются. Это происходит не очень часто и не очень быстро, но в эволюционном масштабе времени размножение интронов, появление новых интронов в эукариотических геномах – это установленный факт. Есть ряд гипотез о механизмах, но точного ответа пока нет.
· Вред, приносимый интронами, как правило, крайне мал (потому что ведь уже есть сплайсосома). Это очень небольшие энергетические затраты на синтез «лишней ДНК» и на сплайсинг. Поэтому давление очищающего отбора, пытающегося избавить геном от интронов, очень слабое.
· Однако этот очищающий отбор может усиливаться, если для организма становится очень важно уменьшить размер своих клеток. Известно, что большие геномы требуют больших клеток – есть положительная корреляция. Поэтому, например, у очень мелких одноклеточных эукариот интронов в среднем, как правило, меньше, чем у крупных (хотя это нестрогая зависимость).
· Прокариоты очень маленькие, и в их геномах очень мало лишнего. Для них энергетические затраты на содержание «эгоистической» ДНК действительно ощутимы, и поэтому очищающий отбор, выбрасывающий из генома все лишнее, довольно силен.
· Поскольку вред от интронов (у эукариот) крайне мал, в ходе эволюции с большой вероятностью в интроне РАНЬШЕ возникает какая-то полезная мутация – например, там появляется полезный новый энхасер или даже маленький полезный ген – чем произойдет и будет поддержана отбором делеция интрона. (Делеция – это мутация, состоящая в выпадении, потере более или менее длинного участка ДНК). Многие «паразитические» или «эгоистические» фрагменты ДНК успевают стать полезными раньше, чем слабенький очищающий отбор успеет от них избавится.
Догмы
Итак, мы вспомнили школьную программу и набрали некий минимум исходных знаний, с которыми мы уже можем начать двигаться вперед. Все остальные «азы» и базовые понятия, которые мы не успели повторить, будем вспоминать и разбирать по мере необходимости, когда они нам понадобятся.

Великие открытия середины прошлого века (расшифровка природы наследственности и изменчивости (ДНК, репликация, генетический код, транскрипция, трансляция, мутации) породили временную эйфорию. В этот период результаты некоторых (действительно великих) открытий превратились в догмы. Очень скоро, однако, выяснилось, что не стоило так торопиться. Любое знание об окружающем мире, любая модель, любая теория, любой вывод – всегда упрощение. Жизнь, как выяснилась, штука очень сложная, хотя она и основана на довольно простых базовых принципах.
Тема ближайших 2-3 занятий – разоблачение догм. Здесь очень важно чувство меры, то есть правильное понимание границ применимости тех или иных научных выводов и обобщений. Все догмы, которые мы будем опровергать, на самом деле верны, но верны лишь в общих чертах, в определенных рамках, не всегда и не везде, из них есть исключения, их нельзя абсолютизировать. Главное – не впадать в крайности.
Вот эти «догмы» - утверждения, которые 40-50 лет назад казались абсолютными истинами, но потом было показано, что они не абсолютны.
· Мутации (изменения последовательности нуклеотидов ДНК) всегда абсолютно случайны
· Однонаправленная передача наследственной информации в ряду ДНК → РНК → Белок (или генотип → фенотип)
· Приобретенные признаки никогда не наследуются
· Принцип «один ген – один белок»
· Эволюция всегда имеет дивергентный характер (ветви эволюционного древа, разделившись, более никогда не сливаются)
· Эволюция основана на безжалостной «борьбе за существование» и конкуренции по принципу «каждый за себя»
Все эти утверждения в значительной мере верны, но не абсолютны. Начнем постепенно разбираться с этими догмами.

«Главная догма»: Однонаправленная передача наследственной информации в ряду ДНК → РНК → Белок (или генотип → фенотип).
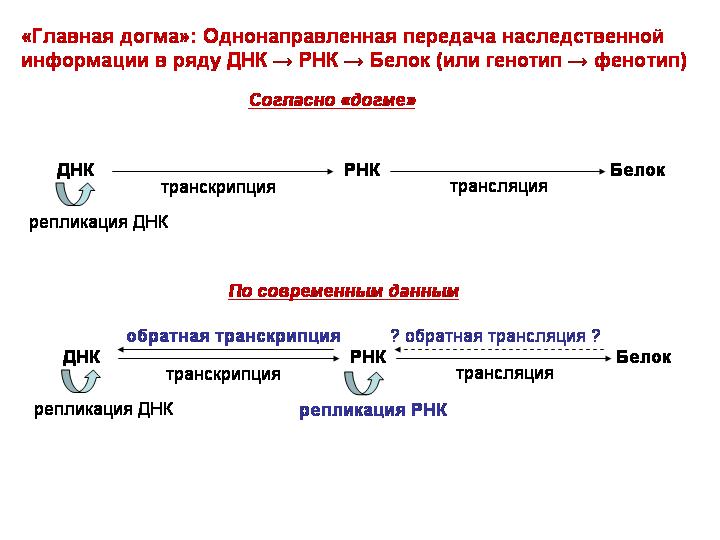
Это правило назвал «центральной догмой молекулярной биологии» сам Фрэнсис Крик (правда, потом раскаялся).
Думали, что информация идет только одним путем. Обратное движение считалось невозможным: то есть у РНК нет никакого способа повлиять на то, что записано в ДНК; у белков нет никакого способа повлиять на то, что записано в РНК и ДНК.
Но потом открыли обратную транскрипцию.

Ее открыли у т.н. ретро-вирусов – это РНК-содержащие вирусы, которые кодируют в своем геноме фермент обратную транскриптазу, она же – РНК зависимая ДНК-полимераза. Этот фермент синтезирует ДНК на матрице РНК, то есть осуществляет обратную транскрипцию. Эта ДНК, синтезированная на матрице вирусной РНК, затем встраивается в геном зараженной клетки и начинает размножаться вместе с ним, как все остальные гены хозяйского генома.
Обратная транскрипция не только в жизни вирусов играет важную роль. Благодаря вирусам, постоянно встраивающимся в геномы живых организмов, гены обратных транкриптаз постоянно попадают в эти геномы. Например, в геноме человека – сотни генов обратных транскриптаз. Это самый распространенный ген в геноме человека.
Это имеет далеко идущие последствия. Обратные транскриптазы могут подвергать обратной транскрипции не только РНК вируса, но и РНК хозяйской клетки. Иногда обратная транкриптаза берет какую-нибудь матричную РНК хозяина, делает из нее ДНК-копию и встраивает в хозяйский геном. Так в наших геномах появляются т.н. ретрогены.
Ретрогены были бы просто дубликатами уже имеющихся в геноме генов (что тоже может иметь важные эволюционные последствия), если бы не одно «но»: обратной транскрипции подвергаются обычно уже зрелые мРНК, прошедшие сплайсинг. Поэтому ретрогены легко отличить от «обычных» генов по отсутствию интронов. А если сплайсинг был альтернативным, то ретроген будет существенно отличаться от своего «прототипа» (в нем не будет некоторых экзонов). Альтернативный сплайсинг – процесс регулируемый, не случайный. Следовательно, здесь открывается потенциальная возможность передачи осмысленной информации от фенотипа к генотипу. Некие крупицы информации о среде и о фенотипе могут в принципе этим путем оказаться записаны в ДНК. Это очень трудный и окольный путь, но он существует.
Пока нет доказанных случаев целенаправленного использования этого пути. Иногда такое происходит случайно (появляются в геномах ретрогены, являющиеся ретрокопиями одного из сплайс-вариантов какого-нибудь исходного гена). Но чтобы клетка делала такое преднамеренно, таких случаев пока не обнаружено. По-видимому, организмы так и не научились использовать эту возможность к собственной выгоде.
Это мы разобрались с обратной транскрипцией.
А как насчет «обратной трансляции»? Так можно было бы назвать процесс синтеза мРНК на матрице белка. Для этого нужны были бы «обратные тРНК», то есть такие молекулы, которые узнавали бы аминокислоту в белке и приделывали к синтезируемой мРНК соответствующий триплет нуклеотидов. В природе такого процесса не обнаружено. Однако искусственно удалось создать «обратные транспортные РНК», то есть рибозимы, которые узнают аминокислоту (аргинин) и приделывают к другой молекуле РНК, которая как бы играет роль мРНК, триплет, который в нормальном генетическом коде соответствует аргинину. Но это – пока лишь занятные упражнения специалистов по рибозимам. Возможно, на заре жизни и существовала обратная трансляция – технически она возможна – но никаких ее следов в современной земной жизни мы не видим.
С другой стороны, обратите внимание, что сам по себе альтернативный сплайсинг – это процесс регулируемого, целенаправленного редактирования наследственной информации, содержащейся в РНК. Некоторым образом это тоже нарушение «главной догмы». Это некий окольный путь, по которому осмысленная информация может передаваться от белков к РНК.

Следующая догма: «Мутации (изменения последовательности нуклеотидов ДНК) всегда абсолютно случайны».
В принципе да, мутации случайны, живым организмам так и не удалось выработать механизм для осмысленного редактирования своих геномов. То есть клетка не может точно рассчитать, какие именно изменения ей нужно в данной ситуации внести в свой геном.
Однако оказалось, что в ходе эволюции у разных организмов появился целый арсенал средств, позволяющих частично контролировать и направлять мутационный процесс (хотя полностью исключить элемент случайности не удалось никому, кроме, возможно, человека).
С одним из таких механизмов мы уже знакомы. У бактерий имеются гены склонных к ошибкам ДНК-полимераз, которые включаются в экстренных случаях. Таким образом, бактерии могут регулировать темп своего мутагенеза. Мутации остаются случайными, но их темп, частота возникновения – оказывается подконтрольной, регулируемой. Тем самым случайность уже загоняется хоть в какие-то рамки. .
Второе исключение из правила о случайности всех изменений ДНК – это процесс создания новых генов из наборов заготовок. Этот процесс происходит в клетках иммунной системы позвоночных животных. Иммунная система позвоночных фантастически эффективна. Она в состоянии по мере необходимости производить новые защитные белки – например, антитела, - способные распознавать и обезвреживать чуть ли не любые чужеродные молекулы (белки, углеводы). В том числе и такие молекулы, которых раньше и в природе-то не было. Например, появился новый вирус. Если он попадет в кровь – через некоторое время там появятся антитела, распознающие и избирательно связывающие поверхностные белки именно этого вируса.
Рассмотрим процесс создания гена антитела в B-лимфоцитах
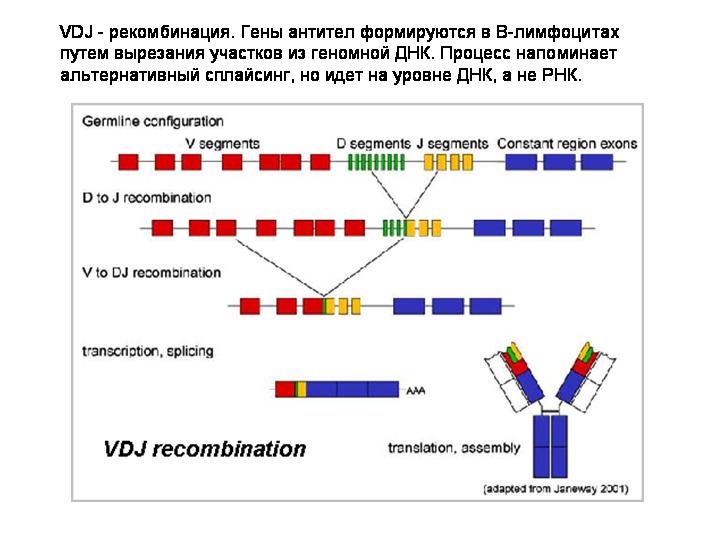
Процесс создания гена антитела из набора заготовок похож на альтернативный сплайсинг, но он осуществляется на уровне ДНК, а не РНК. Это редактирование самого генома, а не матричных РНК.
Какие белки осуществляют эту геномную перестройку в лимфоцитах? Запоминте этот вопрос, мы к нему вернемся чуть позже, когда сможем воспринять и по достоинству оценить ответ :)
За счет комбинаторики формируются B-лимфоциты, продуцирующие огромное множество разных антител. Каждый лимфоцит производит только один тип антител. Соответственно, у него имеется только один ген (точнее, 4). B-лимфоциты размножаются делением, как амебы. Лимфоциты – потомки одного и того же «предка», называются клоном. У каждого клона – свое антитело. На ранних этапах развития организма происходит отбор лимфоцитов на НЕ способность их антител связываться с собственными молекулами организма (чтобы не было аутоиммунных реакций). Те клоны, которые связываются со «своими» антигенами, отбраковываются, остальные – размножаются. В результате получаем большой набор клонов лимфоцитов, которые связываются с чем угодно, только не с собственными молекулами организма.
Это – первый этап. Далее, когда в организм попадает инфекция, осуществляется более точная «подгонка» гена антитела к конкретному антигену.
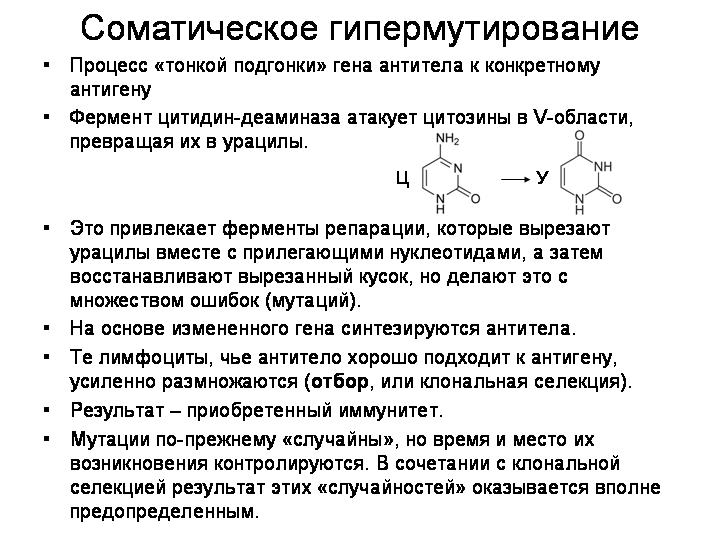

Старинные враги борются друг с другом одним и тем же оружием – при помощи отчасти случайных, отчасти – контролируемых перестроек собственного генома.
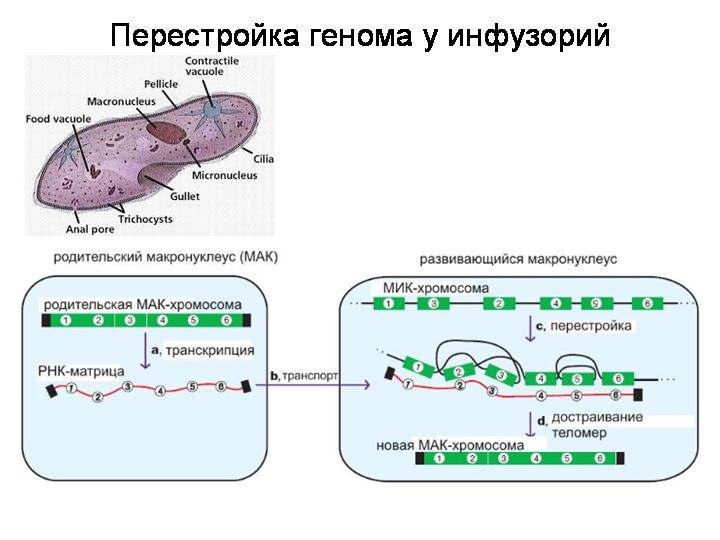
Этот пример описан в книжке.
Процесс тоже похож на сплайсинг, только на уровне ДНК, а не РНК. И еще одно отличие: куски могут меняться местами (при сплайсинге – не могут).
В данном случае особенно интересно то обстоятельство, что в качестве матрицы, направляющей процесс перестройки генома, используются РНК, считанные со «старой» версии генома макронуклеуса. То есть получается, что часть наследственной информации передается у инфузорий с молекулами РНК, а не ДНК.
Еще один вывод: в принципе МОЖНО избавиться от «генетического мусора», как это происходит при сборке МАК-генома. Но клетки этого не делают. И передают при делении потомству МИК-геном со всем содержащимся там «мусором».
Какие белки осуществляют эту геномную перестройку у инфузорий? Запоминте этот вопрос, мы к нему вернемся чуть позже, когда сможем воспринять и по достоинству оценить ответ :)
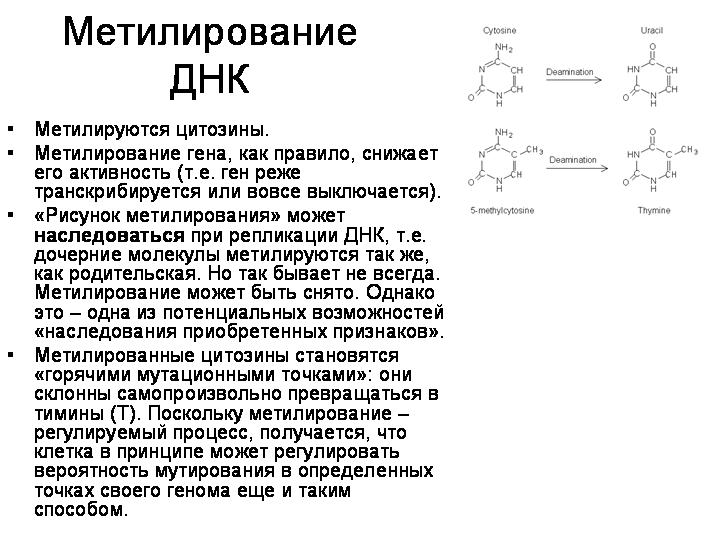
Домашнее задание. Самостоятельная работа: идентифицировать белок по аминокислотной последовательности.
Дата добавления: 2015-06-10; просмотров: 2125;