Если перенос водорода от молекулы донора трудно доказуем, то такие оксидоредуктазы называют редуктазами.
3. Оксидазы – оксидоредуктазы, катализирующие окисление субстратов с молекулярным кислородом в качестве акцептора электронов без включения кислорода в молекулу субстрата.
4. Монооксигеназы – оксидоредуктазы, катализирующие внедрение одного атома кислорода в молекулу субстрата с молекулярным кислородом в качестве донора кислорода.
5. Диоксигеназы – оксидоредуктазы, катализирующие внедрение 2 атомов кислорода в молекулу субстрата с молекулярным кислородом в качестве донора кислорода.
6. Пероксидазы – оксидоредуктазы, катализирующие реакции с пероксидом водорода в качестве акцептора электронов.
Систематическое название образуется: Донор электронов : акцептор электронов – оксидоредуктаза.
Первичные акцепторы водорода окислительно-восстановительных реакций относят к 2 типам дегидрогеназ: никотинамвдзависимым, содержащим в качестве коферментов производные никотиновой кислоты, и флавинзависимым, содержащим производные рибофлавина.
Как уже было сказано, для проявления каталитической активности большинству ферментов необходимо наличие кофермента. Кофермент, локализуясь в каталитическом участке активного центра, принимает непосредственное участие в химической реакции, выступая в качестве акцептора и донора химических группировок, атомов, электронов. Кофермент может быть связан с белковой частью молекулы ковалентными и нековалентными связями. В первом случае он называется простетической группой (пр: FAD, FMN, биотин). Во втором случае кофермент взаимодействует с ферментом только на время химической реакции и может рассматриваться в качестве второго субстрата. Примеры - NAD+, NADP+.
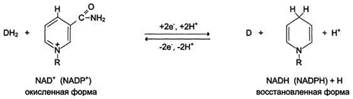 Никотинамидзависимые дегидрогеназы содержат в качестве коферментов NAD+ или NADP+, NAD+ и NADP+ - производные витамина PP. Эти коферменты входят в состав активных центров дегидрогеназ, но могут обратимо диссоциировать из комплекса с апоферментами и включаются в состав фермента в ходе реакции. Субстраты NAD- и NADP-зависимых дегидрогеназ находятся в матриксе митохондрий и в цитозоле. Рабочей частью никотинамидных коферментов служит никотинамид . Большинство дегидрогеназ, поставляющих электроны в ЦПЭ, содержат NAD+. Они катализируют реакции типа: R-CHOH-R1 + NAD+↔ R-CO-R1 + NADH + Н+.
Никотинамидзависимые дегидрогеназы содержат в качестве коферментов NAD+ или NADP+, NAD+ и NADP+ - производные витамина PP. Эти коферменты входят в состав активных центров дегидрогеназ, но могут обратимо диссоциировать из комплекса с апоферментами и включаются в состав фермента в ходе реакции. Субстраты NAD- и NADP-зависимых дегидрогеназ находятся в матриксе митохондрий и в цитозоле. Рабочей частью никотинамидных коферментов служит никотинамид . Большинство дегидрогеназ, поставляющих электроны в ЦПЭ, содержат NAD+. Они катализируют реакции типа: R-CHOH-R1 + NAD+↔ R-CO-R1 + NADH + Н+.
Т.о. NAD+, присоединяя протоны и электроны от различных субстратов, служит главным коллектором энергии окисляемых веществ и главным источником электронов, обладающих высоким энергетическим потенциалом, для ЦПЭ. NADPH не является непосредственным донором электронов в ЦПЭ, а используется почти исключительно в восстановительных биосинтезах. Однако возможно включение электронов с NADPH в ЦПЭ благодаря действию пиридиннуклеотид трансгидрогеназы, катализирующей р-ию: NADPH + NAD+ ↔ NADP+ + NADH.
Флавиновые дегидрогеназы содержат в качестве коферментов FAD или FMN. Эти коферменты образуются в организме человека из витамина В2. Флавиновые коферменты прочно связаны с апоферментами. Рабочей частью FAD и FMN служит изоаллоксазиновая сопряжённая циклическая система. FAD служит акцептором электронов от многих субстратов в реакциях типа: R-CH2-CH2-R1 + Е (FAD) ↔ R-CH=CH-R1 + Е (FADH2), где Е - белковая часть фермента.
 Большинство FAD-зависимых дегидрогеназ - растворимые белки, локализованные в матриксе митохондрий. Исключение составляет сукцинат-дегидрогеназа, находящаяся во внутренней мембране митохондрий. К FMN-содержащим ферментам принадлежит NADH-дегидрогеназа, которая также локализована во внутренней мембране митохондрий; она окисляет NADH, образующийся в митохондриальном матриксе.
Большинство FAD-зависимых дегидрогеназ - растворимые белки, локализованные в матриксе митохондрий. Исключение составляет сукцинат-дегидрогеназа, находящаяся во внутренней мембране митохондрий. К FMN-содержащим ферментам принадлежит NADH-дегидрогеназа, которая также локализована во внутренней мембране митохондрий; она окисляет NADH, образующийся в митохондриальном матриксе.
10. Окислительное декарбоксилирование пирувата и цикл Кребса: последовательность реакций, связь с дыхательной цепью, регуляция, значение.
Окислительное декарбоксилирование пирувата происходит в матриксе митохондрий. Пировиноградная кислота (ПВК, пируват) является продуктом окисления глюкозы и некоторых аминокислот. Ее судьба различна в зависимости от доступности кислорода в клетке. В анаэробных условиях она восстанавливается до молочной кислоты. В аэробных условиях пируват симпортом с ионами Н+, движущимися по протонному градиенту, проникает в митохондрии. Здесь происходит его превращение до уксусной кислоты, переносчиком которой служит коэнзим А.
Превращение пирувата в ацетил-КоА описывают следующим суммарным уравнением:
СН3-СО-СООН + NAD+ + HSKoA → CH3-CO ∼SKoA + NADH + H+ + CO2
Превращение состоит из пяти последовательных реакций, осуществляется мультиферментным комплексом, прикрепленным к внутренней митохондриальной мембране со стороны матрикса. В составе комплекса насчитывают 3 фермента и 5 коферментов:
1) Пируватдегидрогеназа (Е1, ПВК-дегидрогеназа), ее коферментом является тиаминдифосфат (ТДФ), катализирует 1-ю реакцию. 2)Дигидролипоат-ацетилтрансфераза (Е2), ее коферментом является липоевая кислота, катализирует 2-ю и 3-ю реакции. 3)Дигидролипоат-дегидрогеназа (Е3), кофермент – ФАД, катализирует 4-ю и 5-ю реакции.
Помимо указанных коферментов, которые прочно связаны с соответствующими ферментами, в работе комплекса принимают участие коэнзим А и НАД. Суть первых трех реакций сводится к декарбоксилированию пирувата (катализируется пируватдегидрогеназой, Е1), окислению пирувата до ацетила и переносу ацетила на коэнзим А (катализируется дигидролипоамид-ацетилтрансферазой, Е2).
 Оставшиеся 2 реакции необходимы для возвращения липоевой кислоты и ФАД в окисленное состояние (катализируются дигидролипоат-дегидрогеназой, Е3). При этом образуется НАДН.
Оставшиеся 2 реакции необходимы для возвращения липоевой кислоты и ФАД в окисленное состояние (катализируются дигидролипоат-дегидрогеназой, Е3). При этом образуется НАДН.
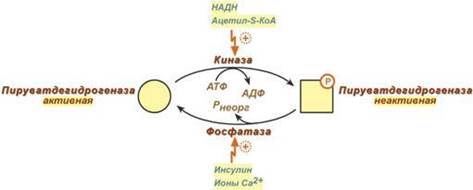
Регулируемымферментом ПВК-дегидрогеназного комплекса является первый фермент – пируватдегидрогеназа (Е1). Два вспомогательных фермента – киназа и фосфатаза обеспечивают регуляцию активности пируватдегидрогеназы путем ее фосфорилирования и дефосфорилирования. Вспомогательный фермент киназа активируется при избытке конечного продукта биологического окисления АТФ и продуктов ПВК-дегидрогеназного комплекса – НАДН и ацетил-S-КоА. Активная киназа фосфорилирует пируватдегидрогеназу, инактивируя ее, в результате первая реакция процесса останавливается. Фермент фосфатаза, активируясь ионами кальция или инсулином, отщепляет фосфат и активирует пируватдегидрогеназу. Таким образом, работа пируватдегидрогеназы подавляется при избытке в митохондрии (в клетке) АТФ и НАДН, что позволяет снизить окисление пирувата и, следовательно, глюкозы в случае когда энергии достаточно. Если АТФ мало или имеется влияние инсулина, то образуется ацетил-SКоА. Последний в зависимости от условий будет направляться либо в цикл трикарбоновых кислот с образованием энергии АТФ, либо на синтез холестерина и жирных кислот.
Образующийся в ПВК-дегидрогеназной реакции ацетил-SКоА далее вступает в цикл трикарбоновых кислот (ЦТК, цикл лимонной кислоты, цикл Кребса). Кроме пирувата, в цикл вовлекаются кетокислоты, поступающие из катаболизма аминокислот или каких-либо иных веществ.
Цикл Кребса протекает в матриксе митохондрий и представляет собой восемь последовательных реакций: связывание ацетила и оксалоацетата (щавелевоуксусной кислоты) с образованием лимонной кислоты, изомеризация лимонной кислоты и последующие реакции окисления с сопутствующим выделением СО2. После восьми реакций цикла вновь образуется оксалоацетат.
Основная роль ЦТК заключается в генерации атомов водорода для работы дыхательной цепи, а именно трех молекул НАДН и одной молекулы ФАДН2. Кроме этого, в ЦТК образуется одна молекула АТФ, сукцинил-SКоА, участвующий в синтезе гема, кетокислоты, являющиеся аналогами АК – α-кетоглутарат для глутаминовой кислоты, оксалоацетат для аспарагиновой.
Главным и основным регуляторомЦТК является оксалоацетат, а точнее его доступность. Наличие оксалоацетата вовлекает в ЦТК ацетил-SКоА и запускает процесс.
Обычно в клетке имеется баланс между образованием ацетил-SКоА (из глюкозы, жирных кислот или аминокислот) и количеством оксалоацетата. Источником оксалоацетата является глюкоза (синтез из пирувата в анаплеротической реакции), поступление из фруктовых кислот самого цикла (яблочной, лимонной), образование из аспарагиновой кислоты в результате трансаминирования.
Примером существенной роли оксалоацетата служит активация синтеза кетоновых тел и кетоацидоз плазмы крови при недостаточном количестве оксалоацетата в печени. Такое состояние наблюдается при инсулинзависимом сахарном диабете, при голодании, алкогольном отравлении или длительной физической нагрузке.
Также некоторые ферменты ЦТК являются чувствительными к аллостерической регуляции метаболитами: Ингибиторы Активаторы
Цитратсинтаза АТФ, цитрат, НАДН, ацил-S-КоА
Изоцитрат-дегидрогеназа АТФ, НАДН АМФ, АДФ
α-Кетоглутарат-дегидрогеназа Сукцинил-S-КоА, НАДН цАМФ
Ацетил-СоА является ключевым промежуточным соединением в превращении всех основных питательных веществ. В аэробных условиях он образуется изсахаров при окислительном декарбоксилировании пировиноградной кислоты, липидов при β-окислении жирных кислот и при гликолитическом распаде глицерина, АК путем образования пирувата (Gly, Ser, Val, Ala, Leu, Ile, Cys), промежуточных соединений цикла лимонной кислоты (Asp, Asn, Glu, Gln, Pro) или др промежуточных соединений (Leu, Ilе, Trp). Внутренняя мембрана митохондрий непроницаема для ацетил-СоА. Поэтому вначале АсСоА конденсируется с оксалоацетатом с образованием цитрата, легко проникающего в цитоплазму. В цитоплазме цитрат распадается под действием цитрат-омыляюшего фермента на оксалоацетат и исходный ацетил-СоА, который используется в синтетических реакциях (синтезе жирных кислот, синтезестероидов и т.д.). Фрагмент молекулы ацетил-СоА, содержащий 2С, вступает в реакцию конденсации с оксалоацетатом (4С), что приводит к образованию цитрата (6С), который в процессе шести последовательных реакций опять превращается в оксалоацетат. Два атома углерода постепенно отщепляются в виде молекул СО2 (одна молекула от ацетил-СоА, другая – от оксалоацетата), а восемь атомов водорода (4 х 2) отщепляются дегидрогеназами, при этом они оказываются связанными с коферментами соответствующих дегидрогеназ. Цикл лимонной кислоты протекает в тесной связи с дыхательной цепью, где большая часть энергии выделяется при окислении восстановленных коферментов. Реакции цикла лимонной кислоты протекают внутри митохондрий, в так называемом матриксе. Ферменты, способные катализировать эти реакции, не связаны с какой-либо биологической структурой. Практически это приводит к тому, что любая отдельная реакция цикла может служить исходным пунктом для дальнейших катаболических или анаболических процессов. Таким образом, цикл лимонной кислоты является одним из нескольких возможных путей превращения ацетил-СоА в организме, направленных на получение наибольшей энергии. Соединения, участвующие в цикле лимонной кислоты, образуют пул промежуточных веществ, дающих начало обратимым процессам в организме. Эти процессы метаболизма связывают в единое целое различные реакции распада и синтеза. С этой точки зрения отдельные реакции цикла лимонной кислоты занимают центральное место в метаболизме. α-Кетоглутарат является самым важным акцептором аминогрупп в реакциях переаминирования. Этот процесс приводит к получению глутамата, из которого могут образоваться глутамин или γ-аминомасляная кислота (последняя может превратиться в полуальдегид яблочной кислоты, который далее окисляется в яблочную кислоту). Глутамин также может выступать в качестве предшественника в синтезе пролина, орнитина, цитруллина, аргинина и других метаболитов.
Оксалоацетат является ключевым соединением для процесса глюконеогенеэа. Из него последова тельными реакциями (через фосфоенолпируват) получаются сахара. В этом случае каждый второй
атом углерода жирных кислот может входить в молекулу сахара. Полный синтез сахаров только
из липидов в организме млекопитающих невозможен, так как при превращениях ацетил-СоА в цикле лимонной кислоты один атом углерода отщепляется в виде CO2. Млекопитающие не способны синтезировать сахара из ацетил-СоА. Обратимое переаминирование оксалоацетата приводит к образованию аспарагиновой кислоты, способной служить исходным соединением для получения пиримидиновых нуклеотидов или некоторых АК. Сукцинил-СоА может образовывать с глицином
δ-аминолевулиновую кислоту, которая конденсируется в порфобилиноген - основное соединение, используемое в синтезе порфириновых структур. Фумаратявляется одним из продуктов разложения ароматических аминокислот, он также образуется из аспарагиновой кислоты в цикле мочевины.
Промежуточные соединения цикла лимонной кислоты включаются в следующие процессы биосинтеза:
а. Глюконеогенез - превращение оксалоацетата в фосфоенолпируват и в сахара.
б. Синтез жирных кислот, который протекает вне митохондрий (ацетил-СоА не может проходить через
мембрану митохондрий, сначала он должен быть превращен в цитрат реакцией с оксалоацетатом; далее цитрат переносится в цитоплазму). Ацетил-СоА вновь образуется в цитоплазме при распаде цитрата с помощью специального АТР-зависимого фермента (цитрат-омыляющий фермент) и затем включается в биосинтез жирных кислот.
в. Взаимопревращения АК. Все реакции начинаются с синтеза трех АК (Ala, Asp, Glu),
получающихся из кетокислот цикла лимонной кислоты за счет переаминирования; центральная роль в этой реакции принадлежит продукту цикла лимонной кислоты - α-кетоглутарату. Аминогруппы различных АК под действием трансаминазы переносятся к α-кетоглутарату. При этом образуется глутамат, а АК, участвующие в реакции переаминирования, превращаются в кетокислоты. Предполагается, что процессы переаминирования протекают преимущественно в цитоплазме. Образующийся в этой реакции глутамат при помощи специфического носителя переносится в митохондрии, где аминогруппа либо отщепляется окислительным дезаминированием, либо ее перенос к оксалоацетату приводит к образованию аспартата. Азот аминогрупп глутаминовой и аспарагиновой кислот в клетках печени оказывается в конце концов в составе мочевины, а образующийся при этом α-кетоглутарат окисляется в цикле лимонной кислоты.
г. Синтез пиримидинов и пуринов. Аспартат образует углеродный скелет пиримидинов. Из глутамина получаются аминогруппы пиримидинов и пуринов.
д. Синтез порфиринов. Сукцинил-СоА является исходным соединением для синтеза порфиринов.
е. Синтез изопреноидов начинается с ацетил-СоА и протекает в цитоплазме. Центральная роль ацетил-СоА в метаболизме определяется тем, что, являясь продуктом катаболизма сахаров, липидов и определенных аминокислот, он может быть или полностью окислен в цикле лимонной кислоты и дыхательной цепи до СО2 и Н2О, или же использован в качестве активного промежуточного соединения для синтеза (его свободная энергия несколько выше, чем свободная энергия АТР).
Дата добавления: 2015-05-26; просмотров: 1450;