Эндотоксины бактерий
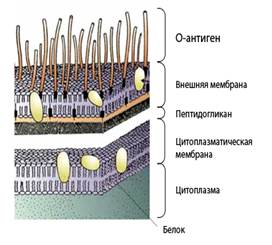
Грамотрицательные бактерииобладают двуслойной клеточной стенкой, которая окружает цитоплазматическую мембрану. Первый слой - очень тонкая (толщиной 1 нм) нелипидная мембрана, состоящая из пептидогликана.Его называют также гликопептидом или мукопептидом. Это сложный матрикс, содержащий полисахаридные цепи, связанные друг с другом поперечными сшивками из коротких пептидных цепей. Второй слой клеточной стенки - липидная мембранатолщиной 7,5 нм. Именно на этой внешней мембране и расположены эндотоксины (липополисахариды). Молекулы эндотоксинаобеспечивают структурную целостность, отвечают за многие физиологические функции, в том числе определяют патогенные и антигенные свойства бактерий.
Структурно молекула эндотоксина делится на три части – Липид А, Кор и О-специфическую цепь (рис. внизу).
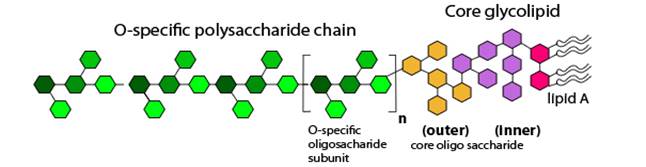
О-специфическая цепь липополисахаридов построена из повторяющихся олигосахаридов. Наиболее распространенными сахарами, входящими в состав О-специфической цепи, являются глюкоза, галактоза, рамноза. Этот участок молекулы придает ей гидрофильные свойства, благодаря которым ЛПС хорошо растворимы в воде. Полисахаридная часть является наиболее вариабельной частью молекулы ЛПC. Часто этот фрагмент молекулы называют О-антигеном, так как именно он отвечает за антигенную активность грамотрицательных бактерий
Кор - центральная часть молекулы, связывающая О-антиген с Липидом А. Формально структура кора подразделяется на внешнюю и внутреннюю части. В состав внутренней части кора обычно входят остатки L-глицеро-О-манногептозы и 2-кето-3-дезоксиоктоновой кислоты (КДО). КДО содержит 8 атомов углерода и в природе практически нигде больше не встречается.
Липид А состоит из дисахарида, фосфата и жирных кислот. Участок Липида А является наиболее константным участком молекулы ЛПС, и его строение схоже у многих бактерий.
• Кроме липополисахаридовв состав внешней стенки грамотрицательных бактерий входят и белки (внешняя мембрана на ¾ состоит из ЛПС, и только ¼ приходится на белковые компоненты). Эти белки вместе с ЛПС образуют белково-липополисахаридные комплексы разного размера и молекулярной массы. Именно эти комплексы и называются бактериальными эндотоксинами. Очищенные препараты, которые используются в качестве стандартов, лишены пептидных фрагментов и представляют собой чистый препарат липополисахарида. Тем не менее, термин «бактериальные эндотоксины» применяется с равным успехом и к естественным эндотоксинам, оказавшимся в растворе в результате разрушения бактерий, и к чистым препаратам ЛПС.
• На внешней стенке одной грамотрицательной бактерии может содержаться до 3,5 млн. молекул ЛПС. После ее гибели все они оказываются в растворе. Эндотоксины грамотрицательных бактерий остаются биологически активными молекулами и после гибели бактерий. Молекула эндотоксина термостабильна и легко выдерживает цикл стерилизации в автоклаве. Малые размеры молекул эндотоксинов позволяют им легко проходить через мембраны, используемые для стерилизации растворов (0,22 мкм). Поэтому эндотоксины могут присутствовать в готовых лекарственных формах, даже произведенных в асептических условиях и прошедших финишную стерилизацию.
• Бактериальные эндотоксины являются исключительно активными (сильными) пирогенами. Для развития лихорадочного приступа достаточно присутствия бактериальных эндотоксинов в инфузионном растворе в концентрации 1 нг/мл (см. http://forums. rusmedserv.com/archive/index.php/t-98927.html). Другие пирогены менее активны, и для развития пирогенного ответа их концентрация должна быть в 100-1000 раз больше. Обычно термины «пирогены» и «эндотоксины» употребляются как синонимы и, хотя не все пирогены являются эндотоксинами, наиболее значимыми являются именно эндотоксины грамотрицательных бактерий.
• Порообразующие токсины. К ним относят бактериальные токсины, функционирующие посредством вставки в плазматическую мембрану хозяина и формирующие в ней трансмембранные поры, приводящие клетку к лизису. Такие токсины еще называют RTX-семейством из-за наличия в их молекулах большого количества повторов [Finlay В., Falkow S., 1997]. Механизм их действия хорошо прослеживается на примере альфа-токсина S.aureus, рассматриваемого как прототип олигомеризующегося пороформирующегося цитотоксина
• Организация и механизм действия токсической молекулы. Большинство токсинов представляют собой А-В структуру. Эта структура предполагает наличие двух компонентов — В-субъединицы, которая участвует в связывании токсина с рецептором на поверхности клетки хозяина и способствует транспортировке токсина в клетку хозяина; и А-субъединицы — проявляющей энзиматическую (токсическую) активность в клетке хозяина. Структура В-доменов зависит от структуры рецепторов-мишеней, с которыми взаимодействует токсин. А-субъединицы более консервативны чем В, особенно в участках, критических для их ферментативной активности
Рис. Механизм действия бактериальных токсинов
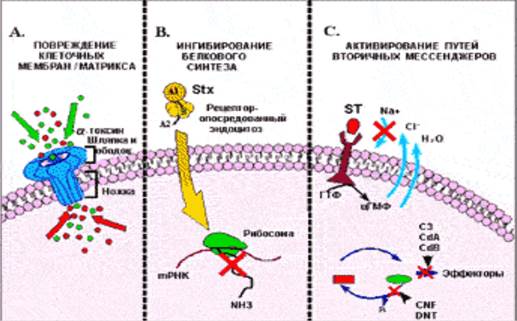
А. Повреждение клеточных мембран альфа-токсином S. aureus.После с ЦПМ клетки, ножка похожего на шампиньон альфа-токсина вставляется в клетку -мишень и вызывает приток, или наоборот, отток из клетки ионов (обозначены как темные и светлые кружки, соответственно). В. Ингибирование белкового синтеза клетки шига-токсином (Stx).Голотоксин, который состоит из энзиматически активной субъединицы (А) входит в клетку через рецептор (Gb3). Затем А-субъединица, обладающая N-гликозидной активностью, отсекает аденозиновый остаток с 28S рибосомальной РНК, что останавливает белковый синтез. С. Примеры бактериальных токсинов, активирующих пути вторичных мессенджеров*. Связывание термостабильного энтеротоксина (ST) с рецептором гуанилатциклазы приводит к увеличению количества ГМФ, который обращает в обратную сторону ток электролитов. Посредством АДФ-рибозилирования или гликозилирования (соответственно), экзоэнзим С3 C. botulinum и токсины A (CdA) и В (CdB) C. difficile, инактивируют небольшие ГТФ-связывающие белки. Цитотоксический некротизирующий фактор (CNF) E. coli и дермонекротический токсин (DNT) рода Bordetella, активируют блокаду эффекторов через дезаминирование .
* ̶ Вторичные посредники (вторичные мессенджеры, англ. second messengers) — это малые сигнальные молекулы, компоненты системы передачи сигнала в клетке.
Дата добавления: 2015-05-21; просмотров: 2306;