Предмет экологии насекомых 5 страница
Заканчивая обзор о влиянии температуры на развитие насекомых, отметим, что с повышением температуры может уменьшаться число линек, и, соответственно, личиночных возрастов.
Влияние температуры на морфологию и окраску
Часто отмечается, что повышение температуры приводит к уменьшению размеров имаго. Это явление может быть связано с отставанием накопления биомассы от метаморфоза. От температуры зависит также частота появления тех или иных форм одного и того же вида. Летом можно найти две формы клопов–водомерок: короткокрылые, которые не способны к миграциям, и активно мигрирующие длиннокрылые. Низкие температуры, при которых длительно сохраняются мелкие водоемы, способствуют появлению короткокрылых водомерок. Наоборот, в жару, когда эти временные водоемы один за другим пересыхают, почти все водомерки оказываются длиннокрылыми и свободно перелетают в поиске благоприятных биотопов.
Окраска многих насекомых в той или иной мере определяется температурой. Так, широко используемые в биологической защите растений осы–бракониды Bracon hebetor Say. при низких температурах темные, почти черные, а при высоких – ярко желтого цвета. Внешне эти температурные формы настолько отличаются друг от друга, что вполне могут быть приняты за разные виды. Правда, между этими крайними формами часто встречаются и переходные, обладающие темными пятнами разных размеров.
Довольно распространены цветовые вариации, вызываемые температурой, у бабочек. Ряд интересных экспериментов с ними провел в начале нашего столетия швейцарский энтомолог М.Штандфусс (1911). С помощью нагрева или охлаждения он, разводя бабочек, пойманных в Швейцарии, смог получить практически все цветовые вариации, свойственные этому виду в пределах от Алжира или Палестины до полярных районов или высокогорий Альп. Так, у бабочки траурницы повышение температуры всегда приводило к уменьшению голубых пятен на крыльях, расширению желтой краевой полосы и повышению яркости и контрастности рисунка. Прикладывая к одной стороне куколки шланг с холодной проточной водой, этот энтомолог получал бабочек, одно крыло которых по окраске походило на южную форму, а другое – на северную.
Температурные вариации описаны не только у имаго насекомых. Например, гусеницы старших возрастов капустной совки Mamestra brassicae L. становятся более темными при низкой температуре (D.Goulson, 1994).
Термопреферендум
Для определения предпочитаемой насекомыми температуры предложено много различных приборов, суть конструкции которых сводится к одному: нагреванию одной части камеры и охлаждению другой – противоположной, т.е. к созданию термоградиента. Самая простая конструкция такого прибора – длинный ящик с дном, сделанным из толстой пластины хорошо проводящего тепло материала, например из латуни. С одного конца пластину нагревают с помощью электрической лампы, а с другого охлаждают льдом. С боковой стороны в ящик вставляют термометры, касающиеся дна, или же, что лучше, к дну приклеивают термопары либо термосопротивления, позволяющие контролировать термоградиент.
Такой прибор имеет ряд существенных недостатков. При большом контрасте температур в его охлаждаемом конце конденсируется капельная влага. Этого можно избежать, или уменьшая температурный контраст, или снижая влажность воздуха во всей камере. Иногда камеру делают из материала, поглощающего влагу, например из гипса. Кроме того, иногда насекомые скапливаются в углах прибора, особенно в холодной зоне, где и оцепеневают. Этого не происходит, если применять круглую или кольцеобразную камеру. Наконец, многие насекомые располагаются на стенках или крышке камеры, что требует особого контроля за температурой этих частей прибора.
По распределению насекомых в камере судят об их термопреферендуме. Оговорим, что подобные данные интерпретировать не так легко. С одной стороны, трудно назвать естественным поведение насекомого, находящегося в такой камере, с другой – скопление насекомых в определенной части камеры может быть связано с уменьшением подвижности при данной температуре, а не с ее предпочтением. Такое торможение активности возможно как в зоне низких температур, так и в зоне высоких. Тем не менее ряд данных по термопреферендуму хорошо соответствует экологии объектов (Б.З. Кауфман, 1995).
Так, живущие на снегу бескрылые комары Chionea предпочитают интервал температур 0...–60° С. Некоторые пещерные ногохвостки скапливаются в зоне с той же температурой, которая имеет место в пещерах (+6...+15° С). Обитающие в лесах виды щетинохвосток рода Machilis имеют оптимум ниже +17° С, а синантропные Lepisma предпочитают зону +15...+20° С. Насекомые, активные летом, всегда предпочитают более высокую температуру, чем весенние или осенние (Т.Е. Россолимо, Л.Б. Рыбалов, 1994). Предпочитаемая температура несколько увеличивается с возрастом имаго.
Интересно, что термопреферендум почти не связан с ареалом вида. Насекомые как северного, так и южного происхождения могут предпочитать одну и ту же температуру (А.Б. Бабенко, 1993). Это подтверждает закон выравнивания среды, согласно которому насекомые ищут в любой географической зоне микроучастки с наиболее подходящим для них микроклиматом.
Термопреферендум меняется, подобно фотопреферендуму, в течение суток. Как правило, насекомые, независимо от образа их жизни, днем предпочитают более высокую температуру, чем ночью. Такие изменения термопреферендума были отмечены у трихограммы, ряда жуков и некоторых других насекомых. Суточные изменения термопреферендума не обязательно связаны с суточными миграциями. Так, у водных личинок комаров Culicidae, круглосуточно находящихся в одной и той же части водоема у поверхности воды, предпочитаемая температура днем выше, чем ночью и утром. Интересно, что личинки Chironomus ночью поднимаются из холодного придонного ила в более теплые поверхностные слои воды. Поэтому ритм их термопреферендума обратен предыдущему и днем они предпочитают более низкие температуры, чем ночью (Б.З.Кауфман, 1980, 1983).
4. ВЛАЖНОСТЬ
Общая характеристика фактора и его измерение
На практике обычно определяют не абсолютную влажность, т.е. количество водяного пара, содержащегося в 1 м  воздуха, а относительную, представляющую собой соотношение реально имеющей место абсолютной влажности (
воздуха, а относительную, представляющую собой соотношение реально имеющей место абсолютной влажности ( 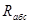 ) и максимально возможной (
) и максимально возможной (  ) при данной температуре:
) при данной температуре:

Иногда используют как параметр влажности дефицит насыщения воздуха влагой, т.е. разность максимальной и абсолютной влажности:
d = 
Влажность чаще всего измеряют психрометром. Этот прибор представляет собой два термометра, причем резервуар одного из них обернут увлажняемой материей. В результате испарения влаги температура этого термометра оказывается ниже температуры воздуха. Сравнивая показатели сухого и влажного термометров, можно с помощью специальной номограммы определить относительную влажность воздуха. Применяются психрометры двух типов: стационарный психрометр Августа и предназначенный для полевых наблюдений психрометр Ассмана. Психрометр Августа имеет емкость с дистиллированной водой, в которой постоянно смачивается конец кусочка батистовой материи, обвертывающего резервуар одного из термометров. Вода из емкости поднимается по материи, как по фитилю. Такие психрометры обычно используются в лабораториях, оранжереях, музеях.
Психрометр Ассмана, в отличие от предыдущего, имеет металлическую изоляцию, защищающую термометры от нагрева в солнечных лучах, а также от механических повреждений. Чтобы термометры, заключенные в металлические трубки, обдувались наружным воздухом, в этом приборе предусмотрен принудительный продув с помощью вентилятора, вращаемого с помощью пружинного механизма. Поэтому перед каждым использованием данный прибор необходимо "заводить", подобно детской игрушке. Этот прибор не имеет емкости с постоянно содержащейся в ней дистиллированной водой и материю "влажного" термометра перед измерением нужно каждый раз смачивать.
Психрометры пригодны только для измерения влажности больших объемов воздуха. Для контроля за влажностью воздуха в небольших экспериментальных камерах часто используют гигрометры, принцип действия которых основан на изменении натяжения легко поглощающего влагу волоска. Эти приборы менее точны, чем психрометры. Для дистанционных же измерений влажности, например в норе или под корой дерева, целесообразно использовать гигросопротивления. При отсутствии фабричных гигросопротивлений их можно изготовить самостоятельно. Для этого берут скрученный из обыкновенной ваты жгут длиной в 5–10 и толщиной 0,5 – 1 см и погружают его в насыщенный раствор поваренной соли, а затем тщательно высушивают. Кристаллы соли хорошо поглощают влагу из воздуха, и, соответственно, электрическое сопротивление жгута будет меняться пропорционально влажности воздуха. Такое гигросопротивление очень инертно, поэтому требуется не менее 2–3 ч для определения влажности.
Перед употреблением гигросопротивления его необходимо градуировать с помощью психрометра или гигрометра.
Для приблизительной оценки влажности также используют кусочки фильтровальной бумаги, пропитанной растворами хлорида кобальта или тиоцианата кобальта. В сухом воздухе эта бумага синего цвета, а во влажном – красная. Влажность воздуха определяют по заранее составленной цветовой шкале.
Влажность воздуха – важнейший фактор, который всегда необходимо контролировать. В лаборатории, не имеющей специальных устройств, влажность воздуха не остается стабильной, а меняется в зависимости от погоды. Любые эксперименты с насекомыми должны проводиться при строго определенной влажности воздуха. Простейший способ создания устойчивой влажности в относительно герметичной камере или термостате поместить в них кювету или другой сосуд с концентрированным раствором той или иной соли. Сосуд должен быть таков, чтобы обеспечить максимально большую поверхность раствора, налитого в него. Для создания более равномерной влажности в камере можно применять небольшой вентилятор, но необходимо убедиться в том, что его работа никак не отражается на поведении подопытных насекомых.
Уровень создаваемой влажности определяется химическим составом соли и температурой. Так, хлорид натрия при температуре 15° создает влажность 78,3%, при 20° – 76,5% , при 25° – 75,9% , при 30° –75,5%. Фруктовый сахар (левулеза) и при 20°, и при 25° создает 55%–ю влажность. Карбонат калия обеспечивает при 20° – 44% влажности, а при 25° – 43%. При тех же температурах хлорид магния создает 35 и 33% влажности. Более подробную таблицу, показывающую влажность воздуха над пересыщенными растворами можно найти в книге И.В.Кожанчикова (1961).
Влияние влажности на насекомых
Скорость испарения влаги насекомым зависит от ее содержания в воздушной среде. Чем больше влажность воздуха, тем дольше сохраняется влага в теле насекомого. Как уже отмечалось, маленькие размеры тела насекомого и, соответственно, относительно большая его поверхность – теоретическая предпосылка для быстрого высыхания. Однако влияние влажности на жизнь насекомого часто не столь очевидно, как влияние температуры или света. Обычно для насекомого оказываются неблагоприятными как слишком низкая, так и слишком высокая влажность, причем эффект влажности определенным образом связан с температурой. Смертность при низкой влажности определяется высыханием, при высокой – прежде всего энтомопатогенными грибами. Для описания совместного влияния влажности и температуры используют так называемые термогигрограммы (рис. 4).
Насекомое не страдает от низкой влажности, если имеет возможность в любое время находить воду для питья, как, например, обитатели берега какого–либо постоянного водоема. То же можно сказать о растительноядных клопах, цикадах, тлях и других насекомых, обладающих хоботком, с помощью которого они прокалывают ткани растения и поглощают их сок.
При жажде насекомые могут компенсировать недостаток воды, поедая любые влажные субстраты. Известно, например, что мигрирующая саранча способна съесть влажные простыни, вывешенные для просушки. При недостатке влаги многие детритофаги могут существенно повреждать сочные части растений.
 Рис.4. Жизнеспособность куколок яблонной плодожорки Laspeyresia pomonella L. в зависимости от температуры и влажности (по В.Шелфорду из В.В.Яхонтова, 1969)
Рис.4. Жизнеспособность куколок яблонной плодожорки Laspeyresia pomonella L. в зависимости от температуры и влажности (по В.Шелфорду из В.В.Яхонтова, 1969)
Надо сказать, что насекомые приспособлены к сохранению влаги настолько хорошо, что практически не прослеживается ожидаемая обратная связь между размерами тела и потерями влаги. М.С.Гиляров (1949) различал три типа приспособлений насекомых к сохранению влаги: морфологические, физиологические и эколого-поведенческие.
К числу морфологических приспособлений относится прежде всего эпикутикула – гидрофобные воскоподобные слои на поверхности покровов. Эпикутикула обычно повреждается в течение жизни насекомого, поэтому имаго с возрастом постепенно уменьшают способность сохранять влагу. Благодаря эпикутикуле потери влаги у насекомых происходят практически только при дыхании. Не случайно, что у многих сухопутных насекомых дыхальца снабжены специальным замыкающим аппаратом. У водных же и почвенных насекомых морфологические приспособления для удержания влаги, за редкими исключениями, отсутствуют.
К числу физиологических приспособлений относятся связывание воды коллоидами, а также поглощение влаги из почвы и даже из воздуха. Так, коксальные мешочки (пузырьки) у щетинохвосток и некоторых других членистоногих непосредственно поглощают воду из влажного субстрата. Всасывание воды может происходить через покровы тела, как это имеет место у личинок жуков–щелкунов. Имеются сведения о способности некоторых насекомых поглощать влагу из воздуха даже при относительной влажности 82 %.
Существенную роль в жизни ряда насекомых играет так называемая метаболическая вода, образующаяся при окислении различных органических веществ, особенно жиров. Метаболизм может быть единственным источником воды для некоторых обитателей пустыни (жуков–чернотелок, питающихся сухими остатками растений), а также обитателей сухих субстратов животного происхождения (жуков–кожеедов и молей). Необходимое для этого повышение интенсивности окислительных процессов требует более активного дыхания, что влечет за собой дополнительную потерю воды. Однако, как отмечает М.С.Гиляров(1970), диффузия паров воды происходит медленнее диффузии газов.
Другой тип физиологических адаптации – это способность переносить высыхание. Обычно насекомые малоустойчивы к нему. Так, личинки некоторых жуков и гусеницы бабочек погибают, когда содержание воды в их теле падает от примерно 75% в норме до 60%. Но личинки некоторых комаров–дергунов, живущие в мелких и быстро пересыхающих лужах на скалах, сохраняют в высушенном состоянии не более 3% воды от массы тела и остаются при этом живыми в таком сухом виде до 10 лет! Высыхание даже необходимо для нормального развития яиц комаров рода Aedes, которые откладывают яйца на края луж.
Эколого–поведенческие приспособления сводятся к активным миграциям в более увлажненные участки в сухое время года, к скоплениям почвенных насекомых на уровне почвенного горизонта, содержащего максимум влаги.
Для определения предпочитаемой влажности (гигропреферендума) используют камеры, в которых создают градиент влажности с помощью растворов различных солей. Над растворами натягивают тонкую газовую сетку, по которой перемещаются насекомые. Результаты этих экспериментов так же сложно интерпретировать, как и при определении термопреферендума. Насекомые предпочитают зоны с влажностью, наиболее соответствующей местам их обитания. Интересно, что у ночных и сумеречных видов, находящихся днем в увлажненных укрытиях, максимальное стремление к влажности наблюдается в дневное время.
В целом насекомые все же довольно чувствительны к уровню влажности воздуха. Отмечается четкая связь между уровнем подвижности многих насекомых и этим фактором. Активность и скорость передвижения максимальны при определенном для каждого вида уровне влажности, причем обычно более резко выражена отрицательная роль низкой влажности. Так, в средней полосе влажность воздуха при ее среднем уровне выше 50% практически не отражается на количестве насекомых, прилетающих на свет. В зоне же пустынь в летнее время ее средний уровень не превышает 20 – 30% и именно она является решающим фактором. Особенно чувствительны к низкой влажности жуки, несколько меньше – бабочки. Саранчовые же почти не реагируют на влажность воздуха.
Если количество прилетающих на свет насекомых может зависеть от влажности воздуха, то время их прилета контролируется почти всегда только освещенностью. Однако эта типичная для географической популяции связь активности с освещенностью меняется в зависимости от среднего уровня влажности воздуха в регионе. Так, лет на свет мелких жуков во влажной зоне Черноморского побережья Кавказа начинается при высокой естественной освещенности, порядка сотен люксов, а в Туркмении – значительно позже в сумерки. В последнем случае жуки как бы дожидаются вечернего повышения влажности. Это справедливо даже для жуков одного и того же вида, обитающих в обоих регионах.
5. ОСАДКИ
Дожди, особенно сильные, могут приводить к гибели некоторых насекомых. Так, ливни смывают с растений значительную часть тлей. Кроме того, во время дождя и после него резко возрастает возможность прилипания насекомых к мокрой поверхности окружающих предметов.
Дождь относительно мало влияет на активность насекомых. Это особенно заметно, если наблюдать лет крупных бабочек на свет. Снижение активности многих дневных насекомых во время дождя объясняется, скорее, уменьшением освещенности. Однако, по–видимому, дождь может и непосредственно подавлять активность особенно мелких насекомых.
 Количество осадков – также важнейшая характеристика климата местности, от которого зависит растительный покров, а следовательно, и формирование населения насекомых. Приблизительную оценку пригодности климата той или иной местности для обитания насекомых можно получить с помощью так называемого, метода климограмм. При построении климограммы по оси ординат откладывают среднюю температуру за каждый месяц, а по оси абсцисс – количество осадков (рис.5). Если последовательно соединить точки на графике, соответствующие каждому месяцу, то возникает фигура, характеризующая климат и его сезонность. Сравнивая климограммы районов, где данный вид может обитать, и тех, где он отсутствует, можно сделать вывод, какие климатические условия необходимы для вида.
Количество осадков – также важнейшая характеристика климата местности, от которого зависит растительный покров, а следовательно, и формирование населения насекомых. Приблизительную оценку пригодности климата той или иной местности для обитания насекомых можно получить с помощью так называемого, метода климограмм. При построении климограммы по оси ординат откладывают среднюю температуру за каждый месяц, а по оси абсцисс – количество осадков (рис.5). Если последовательно соединить точки на графике, соответствующие каждому месяцу, то возникает фигура, характеризующая климат и его сезонность. Сравнивая климограммы районов, где данный вид может обитать, и тех, где он отсутствует, можно сделать вывод, какие климатические условия необходимы для вида.
Аналогичным образом, климограммы, составленные для одной и той же местности за разные годы, можно сравнить с динамикой численности насекомого.
Более компактную, хотя и более поверхностную характеристику климата дает гидротермический коэффициент:

где Ρ – среднегодовое количество осадков, (t° – 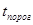 ) – сумма эффективных температур за год (даты с температурой ниже пороговой не учитываются, пороговая температура условно принимается за 6° С). Этот коэффициент пригоден для сравнения климата местностей, лежащих на близких широтах.
) – сумма эффективных температур за год (даты с температурой ниже пороговой не учитываются, пороговая температура условно принимается за 6° С). Этот коэффициент пригоден для сравнения климата местностей, лежащих на близких широтах.
Рис.5.Климограмма г. Бухара (по В.В.Яхонтову, 1969): ось абсцисс – среднее количество осадков за месяц, ось ординат – среднемесячная температура, римскими цифрами обозначены месяцы года.
Однако гидротермический коэффициент может оказаться одинаковым при сравнении климата тундры, где относительно мало осадков и низкие средние температуры, и климата влажных субтропиков, где оба этих показателя велики.
6. АТМОСФЕРНОЕ ДАВЛЕНИЕ
Атмосферное давление в его естественных пределах существенно не отражается на жизнеспособности насекомых, но может заметно влиять на их поведение.
На необычно высокую активность насекомых перед ухудшением погоды, явно имеющую приспособительное значение, уже давно обратили внимание. Но вплоть до настоящего времени вопрос о влиянии атмосферного давления на насекомых остается малоизученным. Большинство авторов сходятся на том, что пониженное давление стимулирует высокую активность лета, питания и спаривания насекомых. По–видимому, пониженное атмосферное давление стимулирует выход бабочек из куколок и вылет веснянок.
Неоднократно отмечалось, что при миграциях насекомые следуют за областями низкого атмосферного давления. Например, массовый лет лугового мотылька явно связан с понижением давления, возникающим при прохождении атмосферного фронта. Очень часто наблюдается концентрация мигрирующей саранчи в областях с низким давлением. Такие скопления, скорее всего, объясняются заносом насекомых ветром, который направлен в зону с низким давлением.
7. ВЕТЕР
Ветер играет особенно важную роль в миграциях насекомых. Возникающие утром в ясную погоду восходящие токи воздуха подхватывают массу взлетающих мелких насекомых и поднимают их на высоту до километра и более. Там эти насекомые с горизонтальными токами воздуха перемещаются на значительные расстояния, а вечером вместе с нисходящими токами воздуха опять опускаются вниз.
Наблюдения за миграциями насекомых показывают, что, чем сильнее ветер, тем больше направление миграции совпадает с направлением ветра. Именно с ветрами связан занос насекомых на очень большие расстояния. Так, однажды на ледниках Шпицбергена было обнаружено значительное количество тлей и цветочных мух. Ближайшая точка, откуда могли прилететь эти насекомые, находится на расстоянии не менее 1300 км от Шпицбергена. Другой пример дальней миграции, облегчаемой ветрами, дает олеандровый бражник Deilephila nerii L., которого обнаруживали в Карелии – на расстоянии в тысячи километров от мест, где может расти олеандр. Этот же бражник регулярно встречается на Черноморском побережье Кавказа, где есть посадки олеандра, но климат не позволяет ему пережить зимнее время. Местная кавказская популяция ежегодно возобновляется в результате залета бабочек, вероятно, из Северной Африки. Этот вид, вообще, отличается исключительно большими способностями к длительным и дальним перелетам.
Скорость ветра зависит от рельефа местности и ее растительного покрова. Под пологом леса она может уменьшаться в 100 и более раз. Даже самые незначительные препятствия типа живых изгородей (ветроломы) способствуют возникновению благоприятных для активности насекомых зон с более спокойным воздухом. В таких зонах скапливается много мелких насекомых: галлиц и других двукрылых, тлей и жуков. Можно предположить, что именно направлениями преобладающих в данной местности ветров объясняется преимущественное заражение вредителями тех или иных участков растительности, особенно леса. Не исключено, что насекомые, готовые к откладке яиц, как бы оседают в зонах наиболее спокойного воздуха, возникающих, например на лесной опушке.
Ветер может существенно влиять и на активность насекомых, особенно на их лет. Большинство насекомых не взлетают при скорости ветра выше определенного уровня. Например, скорость ветра более 3,2 м/с является пределом для полета равнокрылых стрекоз, взлет мелких цикадок невозможен при скорости ветра более 3 м/с, а злаковых мух – больше 1 м/с. На лет более крупных насекомых ветер оказывает меньшее влияние, однако он может существенно нарушать их ориентацию. Даже хорошо летающие крупные бражники перестают кормиться на цветах при ветре более 1,5 м/с. Интересно, что, с другой стороны, в ветреные ночи увеличивается прилет бражников на свет. Не исключено, что ветер стимулирует их миграционную активность.
Ветер заметно влияет на поведение кровососущих двукрылых. Он мешает выходу имаго из куколок, роению, а также нападению на человека. Пороговые скорости ветра, препятствующие нападению комаров, – 2 – 3,5 м/с, мошек и мокрецов – 1 – 2 м/с. При скорости ветра более 4,5 м/ с комары вообще не способны к самостоятельному полету.
Распространение запаха, например полового феромона или пищевого субстрата, существенно зависит от ветра. В силу турбулентности токов воздуха насекомое, ищущее источник запаха, встречает не монотонный градиент концентрации, а чередование плотных струй запаха и чистого воздуха. Поэтому наиболее вероятно, что насекомое ориентируется не по градиенту запаха, а просто летит против ветра, время от времени воспринимая запах, стимулирующий это движение (Л.И.Францевич, 1986).
Интересно резкое воздействие ветра на состав фауны островов и полуостровов, особенно мелких. Так, на полуострове Челекен на Каспийском море сильные ветры уносят в море практически всех летающих насекомых, включая даже синантропных мух. Из-за отсутствия опылителей дикая флора на этом полуострове представлена лишь несколькими видами растений.
8. СИЛА ТЯЖЕСТИ
Гравитационное поле Земли, резко влияющее на формообразование и физиологию крупных животных, не столь существенно отражается на жизни насекомых. У насекомых нет необходимости иметь мощные опорные конечности. Падение с большой высоты обычно не приводит к гибели, так как поверхность их тела настолько относительно велика, что сопротивление воздуха резко замедляет скорость падения. Пострадать от падения на камень или асфальт могут, по–видимому, только крупные жуки и то только в том случае, если они не успеют расправить крылья.
Свободное перемещение насекомых по вертикальной поверхности, поверхности с отрицательным углом и по потолку также связано с их малыми размерами. Сцепления коготков с шероховатостью субстрата, прикрепления с помощью небольших присосок или поверхностного натяжения секрета, выделяемого на конце лапки оказывается вполне достаточным, чтобы удержать тело насекомого от падения.
Некоторые сложности возникают у насекомого в связи с необходимостью удерживать гибкое брюшко над субстратом. Такое поддерживание требует постоянного тонуса мышц, правда, не очень большого, благодаря малому размеру насекомых и, соответственно, высокой эффективности их мускулатуры. Грифельки на брюшке щетинохвосток, по–видимому, представляют собой скользящие опоры, удерживающие брюшко. Нередко напитавшиеся или готовые к откладке яиц насекомые увеличивают свой вес вдвое и даже более. Брюшко у них провисает, поэтому такие насекомые предпочитают садиться на вертикальные поверхности вверх головой. Крупные стрекозы успешно пользуются своим весом, вылезая при линьке из шкурки.
Сила тяжести может не только непосредственно воздействовать на насекомое, но служит и сигнальным фактором. Направление против силы тяжести – это направление к выходу в открытое пространство с меньшей влажностью, большей освещенностью и обычно с более высокой температурой. Кроме того, насекомое, упавшее по каким–либо причинам с растения, возвращается к месту своего обитания по стеблю, используя силу тяжести как ориентир направления.
Такой отрицательный геотропизм у некоторых насекомых может быть очень резко выражен. Известно, что саранчовые некоторых видов, помещенные в садок, поднимаются по стенкам вверх и спускаются к корму, лежащему на дне садка, не раньше чем через двое суток.
Положительный геотропизм используют обитатели почвы. В опытах М.С.Гилярова почвенных насекомых помешали на двухслойные образцы почвы. Если вверху был влажный слой, а внизу совершенно сухой, то более четверти всех насекомых все же спускались в нижний слой и там погибали. В противоположном случае, когда влажным был нижний слой, что обычно и бывает в природе, в нем оказалось 100% выпущенных насекомых.
Геотропизм зависит также от возраста имаго. Например, только что выведшиеся дрозофилы сначала обладают резко выраженным отрицательным геотропизмом, потом он ослабевает, а позже опять усиливается. На протяжении жизни трихограммы каждые 2–3 ч происходит довольно регулярное чередование отрицательного и положительного геотропизма. Отрицательный геотропизм приводит трихограмму на верхушку растения, что способствует взлету и началу миграции с помощью токов воздуха, положительный же геотропизм обеспечивает спуск самки по растению в поисках яиц для заражения. Жертва трихограммы – яйца бабочек – чаще всего находятся на нижней части растения.
Геотропизм меняется и с сезоном. Так, подкоровые клопы Aradidae весной после зимовки стремятся подняться вверх по стволам деревьев, а осенью, наоборот, спускаются вниз. Зиму они проводят у оснований стволов деревьев в местах, хорошо защищенных от холода снегом. Сигналом, приводящим к изменению геотропизма, является устойчивое снижение температуры.
Очень интересен вопрос о приливных воздействиях силы тяжести, связанных с движением Луны вокруг Земли. Приливы имеют место не только на морях и океанах. Твердая земная кора на широте Москвы регулярно поднимается в результате притяжения Луны примерно на 30 см. Вопрос о влиянии силы тяготения Луны был экспериментально изучен швейцарским энтомологом Ф.Шнейдером (F.Schneider, 1964). В опытах регистрировалось направление оси тела майских жуков, находящихся в чашках Петри. Эти наблюдения проводили в подвальном помещении, в темноте и при постоянной температуре. Были обнаружены своеобразные и периодические изменения ориентации жуков. Автор предположил, что одной из причин таких изменений является тяготение Луны. Действительно, когда в опытах поблизости от камер с жуками помещали большой свинцовый груз, создававший локальную силу притяжения, равную лунной, ориентация жуков определенным образом изменялась. Картонные же модели свинцового груза совсем не влияли на ориентацию. Добавим, что жуки были изолированы от груза и не могли его видеть.
Дата добавления: 2015-05-16; просмотров: 1006;