Предмет экологии насекомых 3 страница
Была построена даже действующая модель положительно фототропичного насекомого, имеющая два фотоэлемента и два электромотора, вращавшие независимо друг от друга колеса с правой и с левой стороны модели. Такая модель могла перемещаться ночью по дорожке парка, следуя за человеком, несшим фонарь, и поворачивать за ним по самой запутанной траектории.
Представим себе, что должно произойти с насекомым, у которого один глаз замазан непрозрачным лаком. В результате более высокого тонуса конечностей с одной стороны тела насекомое будет описывать круги на плоскости (манежные движения). В опыте так оно и происходит, что доказывает наличие механизма, соответствующего гипотезе Леба. Однако же через некоторое время насекомое, ослепленное на один глаз, перестает двигаться по кругу и начинает вести себя так, как будто у него сохранились оба глаза. В частности, оно может лететь к источнику света, подобно насекомым с двумя глазами. В другом опыте саранчу помещали в стеклянную трубку, диаметр которой был сходен с диаметром ее тела. При освещении такой саранчи сзади оба глаза были освещены одинаково, но саранча, вместо того чтобы идти вперед, пятилась назад.
 Следовательно, лебовский механизм недостаточен, чтобы объяснить привлечение насекомых светом. Он не позволяет понять, ни почему насекомое, прилетевшее к источнику света, потом покидает освещенную зону, ни особую привлекательность ультрафиолетового излучения, чувствительность глаза насекомого к которому не так уж высока. Кроме того, в природе насекомые летают под разными углами по отношению к Солнцу и Луне и не поворачивают к этим источникам света, как должно было бы быть по представлениям Леба.
Следовательно, лебовский механизм недостаточен, чтобы объяснить привлечение насекомых светом. Он не позволяет понять, ни почему насекомое, прилетевшее к источнику света, потом покидает освещенную зону, ни особую привлекательность ультрафиолетового излучения, чувствительность глаза насекомого к которому не так уж высока. Кроме того, в природе насекомые летают под разными углами по отношению к Солнцу и Луне и не поворачивают к этим источникам света, как должно было бы быть по представлениям Леба.
Другая гипотеза была предложена В.Будденброком (W.von Buddenbrock, 1917). Известно, что у насекомых иногда можно наблюдать менотаксис, т.е. движение под определенным углом к лучам света. Насекомому это легко осуществить, сохраняя изображение источника света в одном определенном омматидии глаза или в их группе. О большом экологическом значении такого типа ориентации будет сказано в дальнейшем. Очевидно, что если насекомое движется, ориентируясь при этом по лучам Солнца или Луны, которые можно считать
параллельными друг другу, то его курс является прямой
линией.
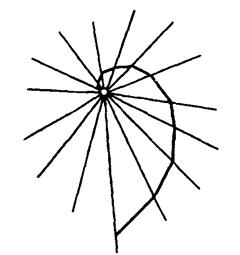
Рис.1 Схематическая траектория летящего на свет насекомого, сохраняющего постоянный угол к лучам света (логарифмическая спираль) (по W. von Buddenbrok из Б.Н.Шванвича, 1949)
Однако при попытке сохранить постоянный угол по отношению к радиально расходящимся лучам искусственного источника света, этот курс неизбежно искривляется, превращаясь в так называемую логарифмическую спираль, неизбежно приводящую насекомое к источнику света (рис.1). Как показывает Г.Н.Горностаев (1984), появление случайных отклонений и поворотов при движении насекомого не меняет окончательного итога.
С этой точки зрения трудно объяснить привлечение насекомых не точечным источником света, а большим экраном, находящимся на стене, почему прилетевшее насекомое может потом улететь от света, а также особую привлекательность ультрафиолетового излучения. Отметим также, что менотаксическая ориентация к лучам Солнца или Луны возникает лишь в особых случаях. Обычно в природе насекомые используют разнообразные наземные ориентиры, обследуют привлекающие их предметы, а их курс представляет собой сложную и довольно беспорядочную кривую.
Очевидно, что механизмы, описанные выше, реально существуют, но включаются только под влиянием определенных мотиваций и представляют собой только части сложного комплекса, называемого - летом на свет. Все эти механизмы имеют в той или иной мере адаптивное значение. Наиболее четкая эколого-поведенческая концепция лета на свет была предложена Г.А.Мазохиным–Поршняковым (1960). По его мнению, свет является универсальным и наиболее надежным индикатором открытого пространства. Здесь особую роль играет ультрафиолетовое излучение, которое практически не отражается наземными предметами, за исключением водной поверхности.
Действительно, случайно залетевшие в комнату дневные или ночные насекомые бьются в стекла окон, пытаясь выйти из замкнутого пространства. Ночью насекомые ведут себя на освещенном экране подобным же образом как на оконном стекле. Попадая ночью в луч света, насекомое начинает воспринимать окружающую его темноту как замкнутое пространство и, стремясь выйти из него, летит к свету.
С этой точки зрения, однако, трудно объяснить, как насекомое, уже прилетевшее к лампе, может опять улететь от нее, почему не происходит подобных нарушений поведения под влиянием лунного света. Кроме того, далеко не все особи одного и того же вида, попадая в освещенное пространство, как-то изменяют свое поведение.
Наиболее вероятно, что лет насекомых на свет представляет собой сложное многоступенчатое явление, причем на каждой ступени работают свои механизмы.
В какой-то мере ступенчатые аналогичные изменения поведения по отношению к свету наблюдал Х.Г.Млетцко (H.G.Mletzko, 1971), выпуская жужелиц на асфальтированную площадку под открытым небом. Сразу же после выпуска жужелица кидается в сторону в любом случайном направлении. Через несколько секунд ее курс приобретает определенную направленность и она бежит либо по направлению к Солнцу, либо от него. Следующий этап – движение жука под определенным углом к солнечным лучам (менотаксис). Такое движение жука почти по прямой линии продолжается до тех пор, пока он не пересекает всю площадку и попадает в более привычную для него обстановку, где можно найти укрытие, пищу или полового партнера. Здесь прямолинейное движение прекращается и жук, не обращая внимания на положение Солнца, начинает руководствоваться другими стимулами.
Кроме того, в естественной обстановке насекомые могут активно перемещаться в поисках зоны с наиболее благоприятным уровнем освещенности. В вечернее время наблюдается массовый вылет насекомых из леса на более открытые места, что хорошо подтверждается сборами оконных ловушек (М.Н.Самков, 1984). Частный случай такого перемещения насекомых в сторону большей освещенности – известный лет насекомых на вечернюю зарю.
По–видимому, аналогичным образом насекомые могут концентрироваться в освещенной зоне около источника света. Однако, это еще не лет на свет, когда наблюдается неудержимое стремление насекомого к лампе. Насекомые, занятые какой–либо деятельностью, например питанием, могут длительно находиться поблизости от горящей лампы, никак не реагируя на нее. Так, по нашим наблюдениям, линейчатые бражники Celerio livornica Esp. на плантациях жасмина даже при низкой сумеречной освещенности порядка 1–0,1 лк продолжали летать над цветами, не реагируя на находившуюся поблизости кварцевую лампу. Подлинный же их лет на свет начинался позже, когда природная освещенность была уже настолько низкой, что они уже не могли находить нектар.
Очевидно, что насекомое в поисках благоприятной для него освещенности ориентируется по градиенту света. Для насекомого же, выходящего из укрытия свет является надежным индикатором выхода в открытое пространство.
После выхода в зону с благоприятной освещенностью насекомое питается, ищет полового партнера, место для откладки яиц и т.д. Однако если насекомое мигрирует, для него наиболее экономичен прямолинейный курс, который легко поддерживать, сохраняя изображение Солнца или Луны в определенных омматидиях глаза. Подобным же образом прямолинейно движутся ползающие насекомые, пересекающие неблагоприятные для их жизни асфальтированные или вытоптанные участки. Иногда этот прямолинейный курс совпадает с направлением дороги и в этом случае можно наблюдать, как жук долго бежит по раскаленному асфальту и не может с него сойти. Интересно, что у лягушек во время их весенней миграции от мест зимовок к водоемам наблюдается подобная же жесткая ориентация по отношению к направлению на Солнце.
Неудивительно, что такое мигрирующее насекомое может избрать в качестве ориентира и искусственный источник света. Тогда оно неизбежно должно приблизиться к нему, причем на достаточном расстоянии от лампы его курс будет относительно близок к прямолинейному и начнет резко искривляться только при приближении к лампе. Как Солнце, так и ночное небо являются источниками ультрафиолетового излучения, поэтому наиболее вероятно, что в качестве ориентира будет избран источник света, излучающий ультрафиолет.
При приближении насекомого на расстояние 30–10 м к источнику света начинается следующий этап поведения, который смело можно назвать патологическим. Дело в том, что если насекомое использует в качестве ориентира естественный источник света, то при движении в его сторону создаваемая им освещенность не меняется. При приближении же насекомого к искусственному источнику света освещенность резко возрастает, особенно на близком расстоянии от лампы. Резко увеличивающаяся интенсивность света буквально ослепляет насекомое. В результате нарушается, ориентация и координация движений, насекомое отклоняется то в одну, то в другую сторону, а иногда из–за нервного шока падает на землю. Конечно, при этом миграционная мотивация может пропадать. Однако насекомое обычно продолжает стремиться к лампе по следующей причине.
Наиболее вероятно, что у ослепленного ярким светом насекомого, глаза которого еще не успели адаптироваться, возникает реакция бегства. Это бегство у хорошо летающих насекомых автоматически направлено в сторону света. Как уже отмечалось выше, такие насекомые при бегстве от любого раздражающего воздействия используют свет как индикатор выхода в открытое пространство. В данном случае эта реакция является парадоксальной, потому что, чем ближе насекомое приближается к источнику света, тем резче раздражающее действие последнего. В результате насекомое продолжает двигаться к лампе, что, в свою очередь, усиливает реакцию бегства (В.Б.Чернышев, 1967, 1973).
Возможно, что поворот назад от лампы для приближающегося насекомого затруднен и тем, что передние омматидии глаза раньше начинают адаптироваться к свету. Тогда то, что находится позади, будет восприниматься как замкнутое пространство. Возникает та же ситуация, которую можно наблюдать на ночном шоссе, когда заяц долго бежит в свете фар автомобиля и не может выскочить из освещенной зоны.
В принципе, реакция бегства может возникать и при неожиданном попадании насекомого в яркий солнечный свет. Однако солнечный свет в земных пределах практически не имеет градиента, поэтому у насекомого движущегося в сторону Солнца, реакция бегства не усиливается и глаза насекомого постепенно адаптируются к свету.
Не исключено, что поблизости от лампы реализуется и лебовский механизм привлечения к свету, основанный на связи тонуса конечностей и уровня освещения глаз.
По-видимому, та же патологическая реакция резкого "притяжения" светом возникает и у некоторых птиц, охваченных миграционным возбуждением. В обычной жизни птиц не привлекает искусственный свет. Однако во время миграций птицы также используют небесные ориентиры.
Наконец, если насекомое приблизилось почти вплотную к лампе и при этом не сгорело или не погибло по какой–либо другой причине, начинается последний этап реакции на искусственный свет. Пока насекомое летает вокруг лампы или бьется в освещенный экран, пытаясь выйти в "открытое пространство", его глаза постепенно адаптируются к свету и общее возбуждение спадает. Нередко ночное насекомое воспринимает яркое искусственное освещение, как наступление дня, и принимает поблизости от лампы позу, характерную для дневного покоя. Но чаще насекомые улетают в темноту, несмотря на то, что их глаза адаптированы к свету. Такое удаление от источника света происходит по гораздо более прямолинейному пути, чем при приближении к нему, причем насекомые летят от света довольно медленно, по-видимому, в связи с необходимостью адаптации глаз к темноте.
Итак, лет на свет особенно типичен для насекомых в силу особенностей работы их нервной системы, включающей значительное количество жестко запрограммированных этапов. В любом случае, лет на свет возникает на основе естественных реакций, имеющих для насекомых большое экологическое значение.
Практическое использование лета насекомых на свет
Источники искусственного света в сочетании с ловушками, собирающими прилетевших насекомых (светоловушками) используются для фаунистических сборов, изучения сезонных и других изменений численности, получения биомассы насекомых в качестве корма другим животным, а также иногда и для уничтожения вредных насекомых (И.А.Терсков, Η.Γ.Κοломиец, 1966).
Сборы насекомых на искусственный свет, особенно с примесью ультрафиолетового излучения, позволяют значительно расширить списки видового состава насекомых, однако некоторые ночные и сумеречные виды совсем не привлекаются светом или прилетают к нему очень редко. Соотношение численности разных видов в сборах на свет, безусловно, ничего не говорит о реальном соотношении их численности в природе. Тем более по этим сборам нельзя судить о реальном соотношении полов в популяции. Так, в Подмосковье на свет прилетают как самцы, так и самки непарного шелкопряда, причем в сборах оказывается вдвое больше самцов, чем самок. На Черноморском же побережье Кавказа на свет прилетают только самцы этого вида. В такой же степени по сборам на свет нельзя судить о физиологическом состоянии самок. Очевидно, что отсутствие в сборах самок того или иного физиологического возраста говорит либо о том, что их еще нет в природе, либо, что они еще или вообще не летят на свет.
Аналогичным образом суточный ритм прилета на свет дает сильно искаженную картину суточной активности. Конечно, если насекомые вообще не летают в данное время суток, трудно ожидать, что они в это время полетят на свет. Тем не менее одновременные наблюдения за уровнем активности бабочек–совок разных видов в садках и за летом таких же бабочек на свет показывает отрицательную корреляцию между общим уровнем активности и численностью сбора на свет (В. Д Кравченко, 1981). Чаще всего насекомые становятся активными существенно раньше, чем наблюдается их лет на свет.
На практике часто используют светоловушки для слежения за численностью насекомых разных видов. Насколько получаемая таким образом информация соответствует истине? Безусловно, нет строгого соответствия между изменениями численности в природе и величиной сбора на свет. Как доказывает сравнение сборов одних и тех же видов насекомых оконными ловушками и светоловушками, первое появление насекомых в сборах на свет может запаздывать на 2–3 недели (М.Н.Самков, 1984). Такое расхождение объясняется тем, что весной и в начале лета эти насекомые летают раньше вечером при таких высоких освещенностях, когда лет на искусственный свет еще невозможен.
Массовый прилет на свет, скорее всего, свидетельствует о высокой численности. Однако известны примеры, когда массовые сборы имели место только за счет пролетающих через данный район мигрирующих насекомых, например бабочки–карадрины Laphygma exigua Hb. И до массового лета на свет, и после него численность карадрины в данном районе была почти равна нулю, а вспышка массового размножения возникла за тысячу километров от места наблюдения. Малый прилет тем более не говорит о низкой численности вида. Улов на свет определяется не только численностью, но также и физиологическим состоянием насекомых, метеорологической и геофизической обстановкой, уровнем лунного освещения. Все это существенно усложняет интерпретацию результатов сборов насекомых на свет. Тем не менее именно светоловушки оказываются во многих случаях наиболее надежным и удобным способом для контроля за численностью вредных видов, а также для выявления карантинных объектов.
Светоловушка, особенно с приспособлением для всасывания воздуха, в благоприятные ночи может собирать килограммы насекомых. Однако нельзя не согласиться с Г.Н.Горностаевым (1984), что рекомендации по использованию собранных на свет насекомых для подкормки домашней птицы и рыбы в прудах целесообразно использовать только в короткое время массового лета тех или иных вредителей, а в целом они не только вредны, но и безнравственны. Применение светоловушек в качестве истребительного средства оказывается в большинстве случаев нерациональным, несмотря на большое количество вылавливаемых насекомых. С одной стороны, светоловушки, подобно инсектицидам, вместе с вредными уничтожают массу редких и полезных насекомых. С другой – процент привлеченных светом насекомых очень мал по сравнению с реальной численностью популяций. Часто на свет прилетают самки насекомых, уже отложившие яйца. Эксперименты показывают, что для успешной защиты урожая на поле требуется расставить значительное количество светоловушек и подвести ко всем электроэнергию. Тем не менее есть ряд сообщений об удачной защите поля, например, от табачного бражника или от кукурузного мотылька, о защите растений закрытого грунта и шампиньонов в теплицах. Так как применение светоловушек довольно дорого и сложно, то их можно рекомендовать только для небольших участков с наиболее ценными сельскохозяйственными растениями.
Разработано много различных конструкций светоловушек. Чаще всего они представляют собой воронку, располагаются под лампой, которая направляет падающих насекомых в контейнер с фиксирующей жидкостью (спирт, формалин, иногда бензин). Если необходимо сохранить насекомых живыми и возможно меньше их повредить, используют большой контейнер (обычно трехлитровую банку), свободно заполненный мятыми полосками бумаги. Сверху ловушка обычно накрыта плоской или слабо большой конической крышкой, предохраняющей от дождя. В ряде конструкций используются также отражающие экраны, поставленные вокруг лампы радиально, так чтобы минимально задерживать свет.
Для южных районов, где обычен массовый лет, наиболее практичной представляется так называемая пенсильванская ловушка. Исходные параметры ловушек этого типа, в том числе отечественной ЭСЛУ–3 следующие: диаметр воронки – 44 см, источник света мощностью 15–30 Вт. В умеренной зоне более стабильные результаты получаются с помощью разработанной Г.Н.Горностаевым модели СКЭЛ (световая, коническая, экранированная ловушка), диаметр которой не менее 60 см, источники света с примесью ультрафиолетового излучения мощностью не менее 200 Вт. Для экспедиционной работы коническая ловушка может быть сделана из полиэтиленовой пленки на складном обруче, что значительно облегчает ее перевозку.
Ранее для сборов насекомых на свет успешно применяли медицинские кварцевые лампы (ПРК–2, ПРК–4, ПРК–8), в последнее время чаще используют лампы уличного освещения типа ДРЛ, дающие более мягкое излучение, не приводящее к ожогам глаз наблюдателя. Впрочем, для увеличения прилета иногда специально разбивают наружную стеклянную оболочку, покрытую люминофором и, таким образом, увеличивают долю ультрафиолетового излучения.
Для фаунистических сборов и коллекционирования целесообразно применять не ловушки, которые обычно портят энтомологический материал, а индивидуальный сбор прилетевших насекомых на освещенном экране. Экран должен быть из бумаги или материи, не обработанной отбеливателем, так как последний поглотает ультрафиолетовые лучи. Лампу при этом целесообразно подвешивать на расстоянии в 50–100 см от экрана, лучше выше головы сборщика. Желательно иметь головной убор, затеняющий лицо от прямого действия света лампы, а также стеклянные или пластмассовые очки. В очках с темными стеклами очень неудобно работать. Следует, однако, помнить, что отраженный от экрана свет также может вызвать ожог глаз и кожи, поэтому лучше располагаться на расстоянии в несколько метров от экрана и подходить к нему по мере необходимости. При ожоге глаз целесообразно применять альбуцид и также крепкий настой чая.
Роль ультрафиолетового излучения в жизни насекомых
В отличие от позвоночных животных и человека насекомые видят ультрафиолетовые лучи, воспринимая их как определенный цвет. Эта особенность зрения насекомых объясняется тем, что покровы их глаза проницаемы для ультрафиолетового излучения в отличие от хрусталика позвоночных. Соответственно на цветках, а также на крыльях насекомых при съемке в ультрафиолетовых лучах в ряде случаев выявляются рисунки, скрытые для глаза человека. Так, основания лепестков и генеративные органы цветка при ультрафиолетовом освещении выглядят несколько темнее, чем остальная его часть, что соответствует обычным для цветков рисункам – указателям местоположения нектара (Г.А.Мазохин–Поршняков, 1965). Рисунок иногда резко проявляется в ультрафиолете и на крыльях бабочек. Например, у наших обычных белянок самки отражают 4–5% ультрафиолетового излучения, а самцы в этом излучении совершенно темные. Самцы бабочек–лимонниц, в отличие от самок, имеют большое ультрафиолетовое пятно на переднем крыле. Интересно, что крылья белых дневных бабочек обычно слабо или совсем не отражают ультрафиолетовые лучи. Ночные же белые бабочки отражают от 20 до 55% этих лучей, что делает их особенно заметными для других насекомых при свете ночного неба (Г.А.Мазохин–Поршняков, 1965).
Зрение насекомых обладает удивительными возможностями. Так, по наблюдениям Бертхольфа (L.M.Bertholf, 1932) глаз мухи–дрозофилы способен воспринимать ультрафиолетовое излучение с длиной волны 253,7 нм. Солнечный свет за пределами земной атмосферы содержит ультрафиолетовые лучи и с меньшей длиной волны, однако такое коротковолновое излучение задерживается верхними ионизированными слоями атмосферы. Нижняя граница для ультрафиолетового излучения на поверхности Земли – около 300 нм.
Однако у поверхности Земли воздух достаточно прозрачен для коротковолнового излучения с меньшей длиной волны. Источниками же такого излучения могут быть делящиеся или отмирающие клетки. По А.Г.Гурвичу (1923), митогенетические лучи наблюдаются в диапазоне от 190 до 325 нм Довольно интенсивное ультрафиолетовое излучение возникает при делении клеток дрожжей, субстраты с которыми ищут самки дрозофил для откладки яиц. Напомним, что именно у дрозофил была обнаружена способность воспринимать коротковолновое излучение. Не исключено, что дрозофила видит развивающиеся клетки дрожжей сверкающими особым ультрафиолетовым светом. Возможно также, что другие насекомые видят точки роста растений, а также поврежденные листья, как излучающие собственный свет.
Как уже отмечалось выше, ультрафиолетовое излучение для насекомого – это прежде всего или цвет неба, или отражение неба водной поверхностью. Движение в сторону просветов неба среди растительности – это выход в открытое пространство, в котором нет препятствий для свободного полета. По–видимому, в этом главная причина особой привлекательности ультрафиолетовых лучей для насекомых, спасающихся от опасности. Когда различных насекомых помещали в камеры, освещенные с двух сторон двумя лучами монохроматического света разных длин волн, то практически всегда оказывалось, что привлекательность ультрафиолетовых лучей в 4–6 раз выше, чем других лучей спектра. Напомним, что чувствительность глаза насекомого к ультрафиолетовым лучам по сравнению с другими областями спектра относительно невысока. Глаз насекомого гораздо более чувствителен к зеленым лучам – обычному для них фону.
В других экспериментах В.Б.Чернышевым (1959) была использована камера, имеющая два окошка, закрываемых разными светофильтрами. Пойманных в поле различных насекомых помещали в эту камеру и наблюдали, в какое окошко они бьются, пытаясь выйти наружу. Опыты проводили тут же в поле, и камера была освещена со стороны окошек солнечным светом. Результаты были те же, что и в описанных выше лабораторных опытах: насекомые особенно привлекались ультрафиолетовым излучением. Такое предпочтение обнаруживалось даже в том случае, когда сравнивалась привлекательность окошек с темным для глаза человека увиолевым фильтром, хорошо пропускающим ультрафиолетовое излучение, и совершенно прозрачным для нашего глаза фильтром, задерживающим только ультрафиолет.
При выборе экранов для сбора насекомых на свет следует помнить об этих особенностях поведения насекомых. Так, отбеливатели тканей, превращающие ультрафиолетовые лучи в видимые для человека, делают эти ткани более светлыми для нашего глаза. Но привлекательность экранов, обработанных отбеливателем, для насекомых заметно снижается.
Роль инфракрасного излучения в жизни насекомых
О возможности восприятия насекомыми инфракрасных лучей известно гораздо меньше. Чувствительность глаза насекомого довольно резко спадает в красной области спектра. Большинство насекомых плохо видит при красном освещении, что часто используется экспериментаторами для ночных наблюдений. Инфракрасное излучение, если оно воспринимается насекомым, должно, скорее всего, действовать не на зрительные, а на какие-то другие рецепторы.
Предполагалось, в частности, что златки и усачи находят в полете зону бывшего лесного пожара, пользуясь тем, что выгоревшая часть леса гораздо больше прогревается солнцем и является мощным источником инфракрасного излучения.
Была выдвинута также гипотеза о роли инфракрасного излучения в сближении полов у ночных бабочек. Температура тела летящей бабочки заметно выше температуры окружающего воздуха и, следовательно, оно является источником инфракрасных лучей. Поток этих лучей, испускаемых телом летящей бабочки, ритмически прерывается с определенной частотой взмахами крыльев. Предполагается, что самец находит и отличает самку, своего вида, воспринимая модулированные таким образом инфракрасные лучи (P.S.Callahan, F.Lee, 1974).
Роль света в пространственной ориентации насекомых
Экологическая роль менотаксиса была отмечена при анализе причин лета насекомых на свет. Такая ориентация под определенным углом к световому потоку дает возможность насекомому, двигаясь прямолинейно, преодолевать максимальное расстояние при миграции, а также быстро пересекать неблагоприятную для него зону. Однако Солнце движется по небосводу, поэтому при сохранении постоянного угла по отношению к солнечным лучам реальный курс насекомого на местности будет постепенно изменяться. Очевидно, что подобный тип ориентации совершенно неприемлем, когда насекомое должно двигаться в строго определенном направлении независимо от времени суток. Чтобы все время сохранять один и тот же курс, некоторые насекомые способны вносить регулярную поправку на движение Солнца по небосводу.
В условиях эксперимента, когда такому насекомому предлагается в качестве ориентира неподвижная горящая лампа, направление его курса должно определенным образом изменяться в течение суток. Именно такая ориентация по небесным телам с поправкой на время – астротаксис – была обнаружена у некоторых живущих на поверхности воды клопов–водомерок, околоводных жуков–стафилинов, а также у пауков, способных передвигаться по поверхности воды. Сходная ориентация имеет место и у ракообразных, живущих у края воды на берегу моря. Все эти членистоногие при необходимости безошибочно возвращаются на берег или в воду, имея заранее строго регламентированное для каждой популяции направление бегства. Интересно, что если прибрежных жуков–стафилинов, обладающих подобной реакцией возвращения на берег, перенести от одного берега реки к противоположному, то эти жуки побегут в совершенно ложном для их спасения направлении – к середине реки.
Очень хорошо изучен астротаксис на примере ориентации медоносной пчелы. Здесь наблюдается запоминание строго определенного курса на местности по направлению к взятку и обратно – к улью. На протяжении жизни пчелы этот курс должен многократно изменяться, чем он отличается от жестко установленного курса более примитивных насекомых. Кроме того, пчела способна "переключаться" с курса, ведущего к взятку, на диаметрально ему противоположный при возвращении назад к улью.
3. ТЕМПЕРАТУРА
Общая характеристика фактора.
Температура – важнейший экологический фактор, особенно резко влияющий на жизнь пойкилотермных животных, к числу которых относятся и насекомые. Температура более или менее закономерно меняется в течение суток и от сезона к сезону. Наиболее резкие суточные изменения температуры отмечены в пустынях. Быстрее всего нагревается в солнечных лучах и остывает после захода Солнца поверхность почвы, если она не затенена растительностью. Чем глубже находится слой под поверхностью почвы, тем меньше там суточные и сезонные колебания температуры и тем ближе она к среднегодичному уровню.
Нагрев поверхности пропорционален синусу ее наклона по отношению к солнечным лучам. Соответственно, на склонах, ориентированных по-разному к странам света, температура воздуха над поверхностью может отличаться на несколько градусов. Как уже отмечалось, растительный покров, особенно лес, существенно сглаживает колебания температуры.
Особые температурные условия возникают у самой поверхности растений. Листья растений и стволы деревьев могут нагреваться в солнечных лучах и быть теплее воздуха на несколько градусов. Однако, если растение находится в пустыне или полупустыне при низкой влажности воздуха и испаряет много воды, подобно бахчевым или хлопчатнику, температура поверхности листьев и стеблей в середине дня оказывается ниже температуры воздуха на 15° и более. Особые условия создаются в нижней части стебля, пропускающего из глубины почвы поток воды к испаряющим листьям. Здесь днем во время интенсивной транспирации температура может быть даже ниже, чем ночью, когда транспирация останавливается. В целом колебания температуры на поверхности растения часто существенно сглажены по сравнению с температурой воздуха.
Своеобразные условия могут возникать под корой деревьев, особенно стоящих на открытых местах. Здесь прогрев солнечными лучами может существенно повышать температуру, особенно в весеннее время при еще малом угле подъема Солнца.
Дата добавления: 2015-05-16; просмотров: 906;