Что происходит при возникновении различий
В 20‑е годы немецкий ученый Фогт сумел проследить судьбу разных частей яйца в развитии. Для этого он окрашивал небольшие участки бластулы амфибий безвредными витальными красителями и наблюдал движение этих участков во время гаструляции и их последующую судьбу. Так была составлена нанесенная на рисунок бластулы карта презумптивных зачатков, т. е. обозначены те клеточные территории, которые при нормальном развитии станут теми или иными зачатками, а потом и органами. Важно подчеркнуть, что презумптивная судьба клеток реализуется только при нормальном развитии. В эксперименте же их судьбу на этой стадии еще нетрудно изменить. Задача биологии развития во многом и заключается в том, чтобы выяснить во всех подробностях, как происходит реализация этой карты. Или, иначе, каким образом расположенные рядом районы, состоящие на стадии бластулы фактически из одинаковых клеток, становятся зачатками разных органов, клетки которых по мере развития все больше отличаются друг от друга.
Эта задача складывается из двух других в значительной степени самостоятельных проблем, хотя на первый взгляд их трудно разграничить. Первую проблему можно сформулировать так: как происходит дифференциация зародыша, т. е. каким образом он распадается на группы неодинаковых клеток и как достигается строго определенное расположение сначала зачатков, а затем и органов развивающегося организма. Вторая проблема состоит в попытке понять, как происходит специализация клеток, т. е. последовательные изменений, превращающие эмбриональную клетку или ее потомков в высокоспециализированные клетки взрослого организма характерного строения и с узко ограниченной функцией.
Оба явления – дифференциацию и специализацию – в принципе можно представить независимо друг от друга. Можно вообразить искусственную ситуацию, когда возникнет правильное расположение зачатков и произойдет морфогенез органов, хотя клетки их внешне почти не будут отличаться друг от друга (в действительности этого никогда не бывает). А можно представить и обратную картину – беспорядочное скопление высокодифференцированных клеток: нервных, мышечных, эпителия кожи и т. д. (нечто похожее встречается в особых опухолях – тератомах).
В этой главе речь идет только о первой проблеме, т. е. о том, как происходит закономерное и прогрессирующее разделение зародыша на все большее число составляющих его зачатков различных органов. Мы будем здесь больше говорить о том, как возникают различия между клетками, и меньше о том, в чем эти различия состоят.
Напомним элементарные основы эмбриологии амфибий. В результате сложных перемещений из верхней (анимальной) части бластулы образуется эктодерма, из нижней (вегетативной) – энтодерма, а из промежуточной, или экваториальной, зоны – мезодерма. В конце гаструляции все эти три зародышевых листка располагаются концентрическими слоями: эктодерма снаружи, энтодерма внутри, мезодерма между ними. Из части эктодермы на спинной стороне зародыша образуется нервная пластинка, которая свертывается в нервную трубку. Передняя ее часть образует головной мозг и глаза, а задняя – спинной мозг. Остальная эктодерма образует в основном эпителий кожи. Из энтодермы развиваются кишечник и его производные – печень, легкие, поджелудочная железа. Мезодерма дифференцируется наиболее сложно: ее центральная часть, которая при гаструляции оказывается на спинной стороне, образует хорду и спинную мускулатуру, далее по обе стороны от хорды располагаются части мезодермы, из которых образуются соединительная ткань, хрящ и кости, еще ниже (ближе к брюшной стороне) возникают почки, а на брюшной стороне – сердце, сосуды и клетки крови. Таким образом, в раннем эмбриогенезе зародыш сначала разделяется на три части, а затем каждая из них тем или иным путем дифференцируется еще на ряд зачатков. На более поздних стадиях каждый из этих зачатков образует органы и дифференцируется еще на ряд тканей и клеточных типов,
У других позвоночных гаструляция происходит иначе, но в итоге создается похожий трехслойный эмбрион, зародышевые листки которого дифференцируются приблизительно так же, как у амфибий. Особенностью позвоночных является то, что уже бластула состоит из тысяч клеток и все зачатки даже в начале дифференцировки состоят из очень многих клеток,
У беспозвоночных, у которых также различают и бластулу и гаструляцию, число клеток обычно меньше и дифференциация зародыша нередко происходит на очень ранних стадиях, когда отдельные зачатки состоят из немногих клеток или даже представлены одним бластомером. Уже на стадии 50–100 клеток такой зародыш представляет собой как бы мозаику, у которой судьба каждой клетки уже точно определена. Поэтому развитие многих беспозвоночных, хотя и не всех, называют мозаичным. Этот термин обычно противопоставляют другому – регуляционное развитие. Мозаичным называют такое развитие, когда различия между частями зародыша с самого начала практически необратимы. При регуляционном развитии на ранних стадиях (бластула – гаструла) судьба клеток еще может быть искусственно изменена. Если ранний зародыш лягушки на стадии двух клеток или на стадии тысячи клеток разделить на две части, то каждая из частей обычно способна образовать целый зародыш, только меньшего размера. Если на стадии бластулы удалить значительную часть клеток эктодермы (т. е. на анимальном полюсе), то зародыш все равно будет вполне пропорционален. Очевидно, что в обоих этих экспериментах многие клетки должны дифференцироваться не в то, чем бы они стали, если бы в их судьбу не вмешался экспериментатор. Иными словами, развитие части (половинки или большей части) может регулироваться до целого. Такой способностью к регуляционному развитию обладают в большей или меньшей степени зародыши амфибий, рыб, иглокожих, млекопитающих и некоторых других групп животных.
Различия мозаичного и регуляционного типов развития в определенном смысле количественные: в первом случае судьба клеток необратимо определяется раньше, чем во втором. Ho эти количественные различия говорят и о качественной разнице: у мозаичных яиц большую роль играют одни механизмы развития, а у регуляционных – другие.
При описании клеточной дифференцировки используют два важных понятия – «компетенция» и «детерминация». Компетенцией называют способность клеток при соответствующих внешних воздействиях дифференцироваться в нескольких (обычно немногих) направлениях. Классическим примером компетенции является способность эктодермы под влиянием зачатка хорды и прилегающей к ней мезодермы (хордомезодермы) дифференцироваться в нервную ткань, а без такого влияния – в эпидермис кожи. Хвост головастика лягушки компетентен к тому, чтобы резорбироваться (рассосаться), но эта компетенция реализуется только под действием гормонов, выделяемых щитовидной железой. Другие части головастика, хотя и получают ту же дозу гормона, не резорбируются, так как имеют другую компетенцию (см. следующую главу). Стволовые клетки крови в костном мозгу или селезенке взрослого организма компетентны к дифференцировке по меньшей мере в трех направлениях: в красные клетки крови – эритроциты, в белые клетки – различные лейкоциты и в мегакариоциты.
В эмбриогенезе компетенция быстро меняется во времени: будущая эктодерма (анимальная часть бластулы) компетентна к дифференцировке почти в любом направлении (тотипотентна), на стадии гаструлы она сохраняет компетенцию к дифференцировке в эпидермис и нервную систему, еще немного позже компетенция к развитию нервной ткани теряется. Компетенция энтодермы, напротив, с самого начала очень узка и ограниченна. В ходе дифференцировки одна компетенция закономерно сменяет другую: собственно каждый этап дифференцировки это приобретение не только некоторых новых свойств, но и новой компетенции. И только достигнув окончательного высокоспециализированного состояния – терминальной дифференцировки, как, например, эритроциты или ороговевающий эпидермис кожи, клетки теряют всякую компетенцию. Молекулярная природа компетенции неизвестна, но, очевидно, она должна заключаться в каком‑то особом состоянии регуляторного аппарата клетки или определенных генов, которые хотя еще не активны, но более других готовы к активации.
Детерминация, пожалуй, более четкое понятие. Это такое состояние клеток, когда они уже (часто необратимо) вступили на путь определенной дифференцировки, но внешне сама дифференцировка еще никак не выражается. Молекулярная природа детерминации также неизвестна, но, вероятно, включение определенных генов – если не первый, то непременный этап всякой детерминации. Широкая компетенция эктодермы не позволяет считать ее окончательно детерминированной, хотя при отсутствии особых влияний она дифференцируется в эпидермис. Напротив, компетенция энтодермы так узка, что ее можно называть детерминированной в направлении развития кишечника. Из этого примера ясно, что детерминация – это сужение компетенции. При описании процессов кроветворения чаще употребляют термин «коммитирование». Коммитированной называют клетку, уже осуществившую выбор направления дифференцировки, например в красные или в белые клетки крови.
Между детерминацией и дифференцировкой не может быть четкой границы. Это видно из самого определения детерминации. Оно говорит лишь о том, что доступными нам сейчас средствами мы не видим в клетках проявлений дифференцировки. Может быть, завтра эти проявления мы уже сможем увидеть, например в виде синтеза определенных мРНК. Считать ли их проявлением дифференцировки или условиться называть дифференцировкой только появление особенностей клетки, начиная с синтеза в ней новых белков, – дело вкуса. Поэтому, наверное, правильнее считать детерминацию не процессом, предшествующим дифференцировке, а ее начальным этапом.
И в заключение еще один круг вопросов. В механике развития часто говорили (и иногда используют эти понятия до сих пор) о самодифференцировке и зависимой дифференцировке. В первом случае предполагается, что клетки способны дифференцироваться в каком‑либо направлении благодаря их внутренней структуре и свойствам. Как мы увидим дальше, это действительно часто бывает в раннем развитии – целые районы зародыша или отдельные бластомеры исходно различаются особенностями цитоплазмы. На более поздних этапах развития представить самодифференцировку труднее: для того чтобы две действительно одинаковые клетки развивались разным путем, необходимо, чтобы одна из них получила извне какое‑то воздействие или информацию, которую не получила другая. Однако и в этом случае, возможно, существует механизм спонтанного вступления на путь дифференцировки, когда отдельные клетки выбирают то или иное направление развития с определенной вероятностью, может быть, под влиянием некоего внутреннего устройства, чем‑то напоминающего рулетку.
Однако основным механизмом возникновения различий между клетками являются локальные внешние воздействия. Такая дифференцировка называется зависимой. Эти воздействия могут быть различными – от совсем простых и неспецифических до высокоспецифических, если их вызывают такие соединения, как белки. В настоящее время нам известны десятки примеров зависимой дифференцировки, но, вероятно, ни в одном случае мы не знаем точного механизма таких воздействий. Поэтому мы можем только предполагать, хотя и с большими основаниями, что все локально действующие факторы, вызывающие дифференцировку, в конце концов приводят к активации (включению) определенного набора генов.
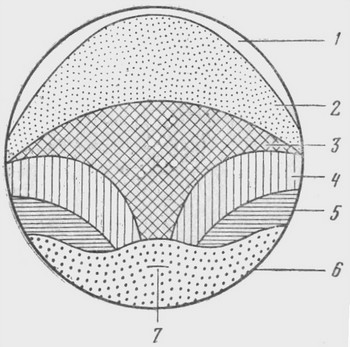
Карта презумптивных зачатков – схема расположения в яйце на стадии ранней гаструлы лягушки материала будущих частей зародыша
Карта могла быть нарисована с большими подробностями, но в действительности на этой стадии между собой различаются только клетки эктодермы, мезодермы и энтодермы. 1 – зачаток эктодермы – будущий эпидермис; 2 – зачаток эктодермы – будущая нервная трубка; 3 – зачаток мезодермы – будущая хорда; 4 – зачаток мезодермы – будущие мышцы спины; 5 – зачаток мезодермы – будущая соединительная ткань, почки, кровь; 6 – зачаток энтодермы – будущий кишечник и его производные (легкие, поджелудочная железа, печень); 7 – бластопор – место, с которого начинается гаструляция: вворачивание энтодермы и мезодермы внутрь зародыша
Дата добавления: 2015-05-08; просмотров: 1037;