Генетический материал бактерий
Бактерии – прокариотические микроорганизмы, генетический материал которых в основном представлен единственной кольцевой двухцепочечной ДНК, называемой генетиками хромосомой. В относительно редких случаях хромосома представлена линейной молекулой ДНК.
 Размер этой ДНК намного превышает размер самой бактериальной клетки. Так, например, у E. coli протяженность хромосомной ДНК равна 1300 мкм (1,3 мм – 4,6 х 106 п.н.), а размер клетки 1,1-1,5 х 2,0-6,0 мкм. Причем ДНК не заполняет всю клетку, а содержится только в ограниченной области, составляющей, весьма приблизительно, одну треть объема клетки.
Размер этой ДНК намного превышает размер самой бактериальной клетки. Так, например, у E. coli протяженность хромосомной ДНК равна 1300 мкм (1,3 мм – 4,6 х 106 п.н.), а размер клетки 1,1-1,5 х 2,0-6,0 мкм. Причем ДНК не заполняет всю клетку, а содержится только в ограниченной области, составляющей, весьма приблизительно, одну треть объема клетки.
|
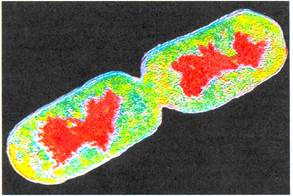
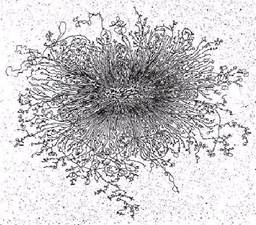
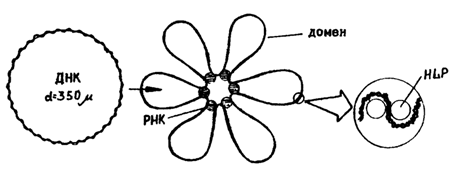
Рис.1. Бактериальный геном и схема уровней его компактизации.
Отсюда следует, что ДНК существует в клетке в высокоупорядоченном (сконденсированном) состоянии в виде компактной структуры. Эта структура, отдаленно напоминая ядра эукариот, получила название нуклеоид и видна в микроскопе только после специфичных для ДНК окрасок (рис.1). В электронном микроскопе она выглядит как образование, состоящее из многочисленных петель, отходящих от плотной центральной области. Образование большого числа (до 140 на геном) петель, называемых доменами, является одним из уровней компактизации ДНК. Каждый домен закреплен у основания молекулой РНК и состоит примерно из 40 т.п.н. ДНК петель находится не в виде свободно вытянутого дуплекса, а имеет второй уровень компактизации за счет скручивания в сверхспиральные образования с помощью связи с белками HLP.
Эти белки имеют небольшой размер, обладают сильноосновными свойствами и прочно связываются с ДНК. По аминокислотному составу они напоминают гистоны эукариот.
Нуклеоид не отделен от цитоплазмы ядерной мембраной и прикреплен к мезосомам – специфическим впячиваниям цитоплазматической мембраны внутрь клетки. Связь ДНК со специфическим участком мембраны необходима для функционирования генома.
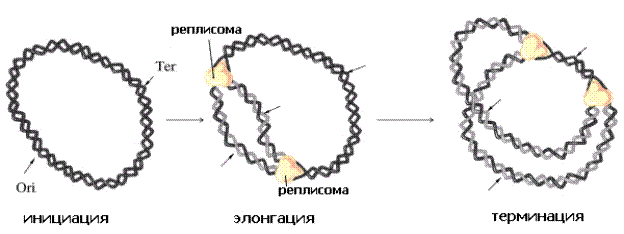
Кольцевая молекула ДНК бактерий (хромосома) представляет самореплицирующуюся генетическую молекулу – репликон. Репликация начинается с точки инициации репликации (ori– orign), локализующейся, как правило, в участке прикрепления ДНК к мембране. От точки инициации репликация происходит последовательно, двунаправленно, по полуконсервативному механизму. Заканчивается репликация в районе терминации репликации (ter), расположенном на участке кольцевой ДНК, противоположном точке начала репликации (рис.2).
|
Рис.2. Репликация бактериальной хромосомы по θ-типу (тета-тип).
Как правило, после репликации следует расхождение копий ДНК к полюсам растущей клетки за счет роста участка мезосомы, расположенного между точками прикрепления дочерних ДНК (рис.3).
Рис. 3. Распределение дочерних копий ДНК и деление клетки бактерий.
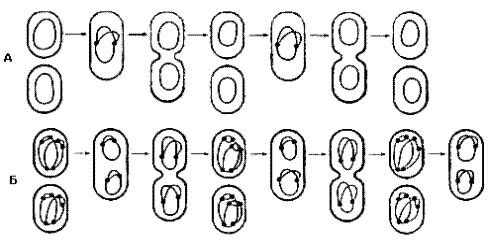
Число хромосом в одной клетке бактерий зависит от стадии развития и физиологических условий роста культуры. В логарифмической стадии роста у E. coli на 1 нуклеоид приходится 2,8 ДНК эквивалентов одного генома, вследствие замедленной сегрегации двух дочерних хромосом, или реинициации новых циклов репликации ДНК еще до деления клетки (рис.4).
|
Рис.4. Число хромосом в клетке в стационарной (А) и логарифмической (Б) стадиях роста культуры.
У некоторых бактерий клетки в норме содержат не одну, а много хромосом. Они могут формировать один или несколько нуклеоидов. Также наблюдается зависимость содержания ДНК в клетке от ее размеров, хотя это не означает соответствующего изменения объема генетической информации.
Для бактериальной ДНК характерна высокая плотность генов (1 ген на 1тпн). ДНК, кодирующая белки, составляет около 85-90% всей ДНК. Средний размер ДНК-последовательностей между генами – только 110—125 п.н. Некодирующая бактериальная ДНК занимает менее 1%, и она обычно представлена в виде транспозонов. Так, в ДНК штамма Escherichia coli K12 линии MG 1655 найдена 41 копия различных транспозонов (IS), которые участвуют в процессах внедрения и исключения плазмид. Многие фаги, исключаясь из генома бактерии не полностью, оставляют там в качестве следа некоторые свои гены. Эти остатки, не способные к самостоятельному перемещению и развитию, называют "криптическими" фагами.
Интроны встречаются в бактериальных геномах крайне редко. Имеются случаи перекрывания генов, где один ген находится внутри другого на той же нити ДНК. Для бактериальных геномов характерны опероны: у Е. coli 27% предсказанных транскрипционных единиц являются оперонами.
В клетке бактерий могут содержаться и другие репликоны, способные существовать отдельно от бактериальной хромосомы. Их называют плазмидами. Плазмиды представляют собой кольцевые (у некоторых видов линейные) молекулы двухцепочечной ДНК различных размеров от 1000 п.н. до почти трети величины самой бактериальной хромосомы. Количество и спектр плазмид в клетках бактерий может варьировать. Часто наблюдаются различия по спектру плазмид даже между клетками разных штаммов одного и того же вида бактерий. Некоторые плазмиды могут встраиваться в бактериальную хромосому, составляя при этом часть репликона бактерии, и могут вновь исключаться из нее, восстанавливая форму автономного репликона. Такие плазмиды называют эписомами.
В генетический материал бактерий могут быть включены и профаги.
Дата добавления: 2015-02-07; просмотров: 7803;