ДВИГАТЕЛЬНЫЙ АНАЛИЗАТОР
Двигательный анализатор осуществляет анализ и синтез раздражений, поступающих из двигательного аппарата, обеспечивает статику и динамику растущего организма ребенка, подвижность и трудовую деятельность человека.
Афферентный отдел его состоит из 3-х нейронов (рис. 11). Тела первых нейронов, псевдоуниполярных клеток, заложены в спинномозговых узлах и узлах V, IX и Х пар черепных нервов.
Дендриты первых нейронов, заложенных в спинномозговых узлах, направляются в составе периферических нервов к костям, суставам и мышцам, заканчиваясь в них рецепторами (проприорецепторами), воспринимающими раздражения, возникающие в связи с изменениями напряжения или растяжения мускулатуры, натяжения или расслабления сухожилий, суставных сумок и т.д. (мышечно-суставное чувство).
Воспринятое раздражение по дендритам передается телам клеток, аксоны которых в составе заднего корешка вступают в спинной мозг и, не прерываясь в сером веществе, поднимаются в заднем канатике. При этом аксоны первых нейронов, воспринимающих раздражение от мышц и суставов нижних конечностей и нижних отделов туловища, образуют нежные пучки, fasciculi gracilis, формирующиеся на протяжении 19 нижних сегментов спинного мозга.
Аксоны клеток, воспринимающих раздражения из мышц и суставов верхних конечностей, верхних отделов туловища и частично шеи, образуют расположенные латерально от предыдущих клиновидные пучки, fasciculi cuneati, формирующиеся на протяжении 12 верхних сегментов спинного мозга. Таким образом, ниже 4 грудного сегмента в заднем канатике проходит только пучок Голля, а выше - оба пучка (Голля и Бурдаха). Из спинного мозга эти пучки поднимаются в продолговатый мозг и заканчиваются в своих ядрах, nuclei fasciculi gracilis et cuneati. Клетки ядер нежного (Голля) и клиновидного (Бурдаха) пучков являются вторыми нейронами. Аксоны их идут двумя путями: меньшая часть, под названием наружных дугообразных волокон, следует через нижние ножки мозжечка к ядру шатра, nucleus fastigii, а большая часть их дугообразно огибает серое вещество, окружающее центральный канал (внутренние дугообразные волокна), переходит между оливами на противоположную сторону и поднимается по бокам от срединной линии кзади от пирамид, получая название медиальной петли, lemniscus medialis. При этом волокна ядер нежного пучка занимают более дорзальное положение. В мосту волокна медиальной петли проходят на границе основания и покрышки, пересекая поперечно идущие волокна трапециевидного тела. В верхнем отделе моста к ним присоединяется спиноталамический путь и весь комплекс волокон поднимается в покрышку ножек мозга, проходя кнаружи и дорзальнее красного ядра. На пути волокна медиальной петли отдают коллатерали к ядрам ретикулярной формации. Заканчивается медиальная петля в вентральном и латеральном ядрах зрительного бугра. Клетки вентрального и латерального ядер зрительного бугра являются третьими нейронами. Аксоны их, составляя таламокортикальный тракт, следуют через заднюю треть заднего бедра внутренней капсулы, позади пирамидного пути, в кору верхних двух третей передней центральной извилины, где заложена ядерная часть коркового конца двигательного анализатора. Эта часть афферентного отдела двигательного анализатора проводит проприоцептивную чувствительность от мышц, сухожилий, костей и суставов конечностей, туловища и частично шеи.


| Nucl.gracilis
(II нейрон)
| |


| Gangl.spinale Th5-Co1
(I нейрон)
| |
| Gangl.spinale
C1-Th4
(I нейрон)
| |
| Nucl. cuneatus
(II нейрон)
| |
| Nucl.tr.solitarii (II нейрон)
| |

| Gangl.rigeminale
(I нейрон)
| |


| Nucl.tr.mesencephalici n.trigemini (II нейрон)
| |



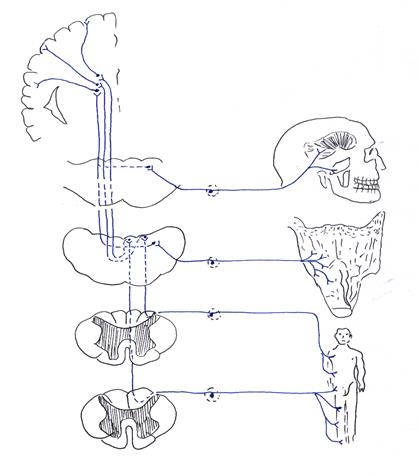 Рис. 11. Пути сомато-сенсороного анализатора
Рис. 11. Пути сомато-сенсороного анализатора
От мышц головы и мышц шеи висцерального происхождения проприоцептивная чувствительность проводится по черепным нервам. Первые нейроны этой части пути двигательного анализатора заложены в узлах V, IX и Х пар черепных нервов. Аксоны их следуют в составе нервов к мезенцефалическому ядру тройничного нерва, nucleus tractus mesencephali nervi trigemeni, и ядру одиночного пучка, nucleus tractus solitarii.
В этих ядрах заложены тела вторых нейронов. Аксоны вторых нейронов переходят на противоположную сторону и присоединяются к медиальной петле, достигая медиального и заднего отделов вентрального ядра зрительного бугра. Клетки этого ядра являются третьими нейронами, аксоны которых следуют в составе таламокортикального тракта в кору нижней трети передней центральной извилины. Таким образом, медиальная петля есть комплекс аксонов клеток ядер нежного и клиновидного пучков, собственного ядра заднего рога, желатинозной субстанции, а также чувствительных ядер тройничного, языкоглоточного и блуждающего нервов. Получив свое начало в продолговатом мозге, она заканчивается в зрительном бугре, являясь проводником общей чувствительности. Необходимо отметить, что часть аксонов первых нейронов (клеток узлов V, IX и Х пар черепных нервов) без перерыва в ядрах стволовой части мозга достигают коры передней центральной извилины. Таким образом, первые нейроны пути двигательного анализатора заложены в спинномозговых узлах и узлах V IX и Х пар черепных нервов, вторые нейроны — в ядрах нежного и клиновидного пучков, а также в чувствительных ядрах тройничного, языкоглоточного и блуждающего нервов, а третьи нейроны — в вентральном и латеральном ядрах зрительного бугра.
Поражение афферентной части двигательного анализатора ведет к выпадению мышечно-суставной (проприоцептивной) чувствительности, что выражается в нарушении представления о положении частей тела в пространстве, ощущения позы и производимых движений, нарушается координация движений (сенсорная атаксия). Больной с закрытыми глазами не может определить положение конечности или её частей, направления и объёма движений в том или ином суставе, движения теряют свою плавность, становятся неправильными, неточными.
Переработанная и проверенная в результате аналитико-синтетической деятельности корковых нейронов информация передается гигантским пирамидным клеткам Беца и пирамидным клеткам V и VI слоев коры, которые дают начало исполнительной системе, обеспечивающей движение импульсов из коры к мотонейронам спинного мозга и двигательным ядрам черепных нервов для осуществления рабочих, исполнительных актов, т. е. ответной реакции на воспринятые раздражения. Эта исполнительная система представлена пирамидным и экстрапирамидными путями или системами.
Пирамидная система осуществляет произвольные, высокодифференцированные движения, посылая из коры полушарий большого мозга через двигательные ядра черепных нервов и α-мотонейроны передних рогов спинного мозга импульсы к мышцам. Она имеет двух- или трехнейронный тип строения и представлена центральными (корковыми), промежуточными (возбуждающими или тормозными) и периферическими альфа-мотонейронами (Рис.12)..
Центральными (первыми) нейронами являются гигантские пирамидные клетки Беца (140 мкм) и большие пирамидные клетки пятого слоя коры передней центральной извилины, gyrus precentralis, лобной доли и околоцентральной дольки, lobulus paracentralis, а также большие и средние пирамидные клетки лобной, теменной, затылочной и височной долей.
Аксоны центральных нейронов, расположенных в пятом слое коры передней центральной извилины и околоцентральной дольки, идут более компактно, нежели аксоны клеток коры других долей. В связи с этим в пирамидной системе выделяют концентрированно расположенные и рассеянные волокна.
Концентрированно расположенные волокна осуществляют целенаправленную ответную реакцию на раздражения, полученные по афферентной части двигательного анализатора, а рассеянные волокна, по-видимому, обеспечивают ответную целенаправленную реакцию на раздражения, полученные по афферентным путям других анализаторов.
Возникнув в различных отделах коры, все эти волокна образуют лучистый венец и, постепенно сближаясь, собираются в компактный пучок, проходящий через колено и передние две трети заднего бедра внутренней капсулы.
В зависимости от ядер, к которым они направляются, в пирамидной системе различают корково-ядерный и корково-спинномозговой пути (рис. 12).
Корково-ядерный путь, tractus corticonuclearis, связывает кору большого мозга со всеми двигательными ядрами черепных нервов. Первыми нейронами этого пути являются гигантские и большие пирамидные клетки пятого слоя коры нижней трети передней центральной извилины. Аксоны их, постепенно сближаясь, образуют компактный пучок, который проходит через колено внутренней капсулы в основание ножки мозга. Здесь он, располагаясь латеральнее от лобно-мостового пути, tractus frontopontinus, отдает часть волокон к ядрам глазодвигательного, nucleus nervi oculomotorii, и блокового, nucleus nervi trochlearii, нервов своей и противоположной стороны. Из основания ножки мозга этот путь спускается в основание моста, отдавая часть волокон к двигательным ядрам тройничного, nucleus motorius nervi trigemini, отводящего, nucleus nervi abducentis, и верхней части ядра лицевого, nucleus nervi facialis, нервов своей и противоположной стороны, к нижней части ядра лицевого нерва только противоположной стороны и коллатерали к собственным ядрам моста, nuclei proprii pontis. Оставшаяся сравнительно небольшая часть волокон спускается в продолговатый мозг и заканчивается в двигательных ядрах IX, Х и XI пар черепных нервов своей и противоположной стороны, в ядре XII пары противоположной стороны. Необходимо подчеркнуть, что не все аксоны первых нейронов вступают в контакт непосредственно с клетками двигательных ядер. Большая часть их образует синаптическую связь с промежуточными нейронами - клетками ретикулярной формации, а последние, в свою очередь, - с клетками двигательных ядер.
Клетки двигательных ядер черепных нервов являются вторыми или третьими нейронами корково-ядерного пути. Аксоны их направляются в составе соответствующих нервов к мышцам головы и частично шеи, а также к мышцам глотки и гортани. Так, в составе глазодвигательного нерва - к внутренней, верхней, нижней прямым и нижней косой мышцам глаза, а также мышце, поднимающей верхнее веко; в составе блокового нерва - к верхней косой, а отводящего - к латеральной прямой мышцам глаза. Аксоны клеток двигательного ядра тройничного нерва идут в составе его третьей ветви ко всем жевательным, челюстно-подъязычной мышце, к мышцам, напрягающим мягкое небо и барабанную перепонку, и переднему брюшку двубрюшной мышцы. В составе лицевого нерва волокна идут к мимическим, шилоподъязычной мышцам и подкожной мышце шеи, а также к мышце стремени и заднему брюшку двубрюшной мышцы. В составе остальных нервов: IX- к шилоглоточной мышце; Х - к мышцам глотки, мягкого неба, гортани и верхней части пищевода; XI- к трапециевидной и грудино-ключично-сосцевидной мышцам; XII- к мышцам языка и мышцам, лежащим ниже подъязычной кости.


| Nucl.n.glosso-pharyngeus
et vagus
| |



| Tr. corticospinalis anterior
| |




| Tr. Corticospinalis lateralis
| |
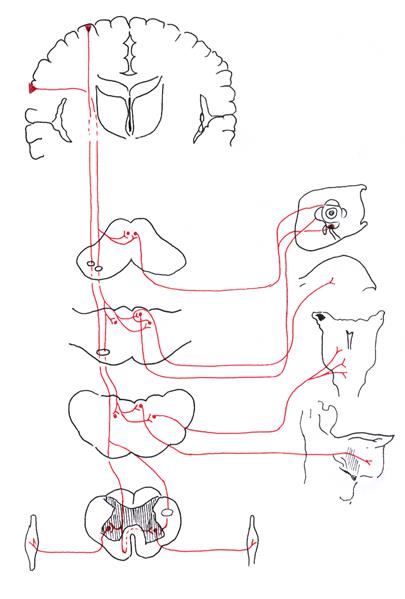
Рис. 12. Пирамидные пути.
В связи с тем, что большинство двигательных ядер черепных нервов (III, IV, V, VI, IX, X, XI пар и верхней части ядра лицевого нерва) получают волокна из коры обоих полушарий большого мозга, одностороннее поражение первых (центральных) нейронов корково-ядерного пути ведет лишь к ограничению произвольных движений, снижению силы мышц, иннервируемых этими нервами, т.е. неполной утрате движений, называемой парезом. Поражение же первых нейронов, аксоны которых следуют к ядру подъязычного и нижней части ядра лицевого нервов из коры только противоположного полушария, ведет к возникновению центрального (спастического) паралича мышц языка и мимических мышц нижнего отдела лица на стороне, противоположной очагу поражения.
Центральный паралич характеризуется резким повышением тонуса мышц при пассивном движении (мышечная гипертония) и сухожильных рефлексов (гиперрефлексия), появлением сопутствующих движений (синкинезия) и патологических рефлексов. Развитие этих симптомов объясняется тем, что при поражении центральных (корковых) нейронов, оказывающих в обычных условиях тормозящее влияние на периферические нейроны, сохраняется целостность рефлекторных дуг, по которым периферические нейроны (клетки двигательных ядер) непрерывно получают раздражения с периферии и, в свою очередь, посылают импульсы к мышцам, повышая их тонус (мышцы напряжены, плотны на ощупь), усиливая сухожильные рефлексы, вызывая появление патологических рефлексов.
Поражение периферических нейронов корково-ядерного пути ведет к возникновению периферического, вялого, атрофического паралича мышц на своей стороне, характеризующегося, вследствие перерыва рефлекторной дуги, отсутствием рефлексов и тонуса мышц с последующей их дегенеративной атрофией.
Корково-спинномозговой путь, tractus corticospinalis, связывает кору больших полушарий с двигательными ядрами передних рогов спинного мозга. Первыми нейронами его являются гигантские клетки и большие пирамидные клетки коры верхних двух третей передней центральной извилины и околоцентральной дольки. Аксоны их следуют в составе лучистого венца и в области внутренней капсулы образуют компактный пучок, занимающий передние две трети ее заднего бедра. Из внутренней капсулы волокна корково-спинномозгового пути спускаются в основание ножек мозга, располагаясь латеральнее от корково-ядерного пути. Здесь весь пирамидный путь занимает центральный отдел основания ножки мозга, составляя 3/5 её. Далее этот путь спускается в основание моста, где поперечными волокнами, возникающими из его собственных ядер и идущими в мозжечок, разбивается на изолированные пучки, хорошо видимые на поперечных срезах моста. Здесь от пирамидных волокон отходят коллатерали к собственным ядрам моста, аксоны клеток которых переходят на противоположную сторону и в составе средних ножек мозжечка следуют к коре полушарий мозжечка .
В продолговатом мозге волокна корково-спинномозгового пути вновь собираются в компактные пучки, расположенные по бокам передней срединной щели - пирамиды. От них отделяется небольшой пучок волокон, идущих через нижние ножки в кору полушарий мозжечка (рис. 13).
Эти волокна вместе с коллатералями, отходящими от пирамидного пути к собственным ядрам моста, обеспечивают мозжечковую поправку к основному импульсу, идущему по пирамидному пути, заключающуюся в преодолении инерции движения.
На границе со спинным мозгом большая часть (80%) волокон пирамид переходит на противоположную сторону и спускается в боковом канатике спинного мозга под названием бокового корково-спинномозгового пути, tractus corticospinalis lateralis, посегментно заканчиваясь в двигательных ядрах передних рогов той же стороны.


Рис. 13. 1 - Пирамидный путь. 2 - Пучок волокон к мозжечку.
Меньшая часть (20%) волокон, под именем переднего корково-спинномозгового пути, tractus corticospinalis anterior, спускается в переднем канатике своей стороны до нижних грудных сегментов, посегментно заканчиваясь в двигательных ядрах передних рогов своей и противоположной стороны. Здесь волокна обоих путей вступают в контакт (синапс) с α-мотонейронами непосредственно, или через посредство промежуточных нейронов (интернейронов). Таким образом, α-мотонейроны являются вторыми или третьими нейронами корково-спинномозгового пути. Аксоны их выходят из спинного мозга через передние корешки и в составе ветвей спинномозговых нервов достигают мышц.
Одностороннее поражение центральных (первых) нейронов корково-спинномозгового пути ведёт к возникновению центрального, спастического паралича мышц конечностей противоположной стороны, при сохранении или ослаблении функции мышц шеи, туловища и промежности. Последнее объясняется тем, что эти мышцы получают двухстороннюю корковую иннервацию. Поражение пирамидного пути в спинном мозге выше шейного утолщения вызывает паралич мышц верхней и нижней конечностей одноименной стороны, а ниже — мышц нижней конечности той же стороны.
Поражение периферических двигательных нейронов (клеток передних рогов и их аксонов) ведет к возникновению периферического, вялого или атрофического паралича мышц той же стороны, характеризующегося утратой рефлексов, потерей тонуса мышц с последующей их дегенеративной атрофией.
Дата добавления: 2015-02-07; просмотров: 4857;