брожение
«Брожение» - это сугубо микробиологический термин. Он характеризует энергетическую сторону способа существования нескольких групп эубактерий, при котором они осуществляют в анаэробных условиях окислительно-восстановительные превращения органических соединений, сопровождающиеся выходом энергии, которую эти организмы используют. Поскольку брожение протекает без участия молекулярного кислорода, все окислительно-восстановительные превращения субстрата происходят за счет его «внутренних» возможностей. Процесс брожения связан с такими перестройками органических молекул субстрата, в результате которых на окислительных этапах процесса высвобождается часть свободной энергии, заключенной в молекуле субстрата, и происходит ее запасание в молекулах АТФ. В процессе брожения, как правило, происходит расщепление углеродного скелета молекулы субстрата.
Круг органических соединений, которые могут сбраживаться, довольно широк. Это углеводы, спирты, органические кислоты, аминокислоты, пурины, пиримидины. Химическое вещество может быть подвергнуто сбраживанию, если оно содержит неполностью окисленные (или восстановленные) углеродные атомы. В этом случае есть возможность для окислительно-восстановительных преобразований между молекулами (или внутри одного вида молекул), возникающими из субстрата. В результате одна часть продуктов брожения будет более восстановленной, другая - более окисленной по сравнению с субстратом. Продуктами брожений являются различные органические кислоты (молочная, масляная, уксусная, муравьиная), спирты (этиловый, бутиловый, пропиловый), ацетон, а также CO2 и H2. Обычно в процессе брожения образуется несколько продуктов. В зависимости от того, какой основной продукт накапливается в среде, различают молочнокислое, спиртовое, маслянокислое, пропионовокислое и другие виды брожений.
Следовательно, в каждом виде брожения можно выделить две стороны: окислительную и восстановительную. Процессы окисления сводятся к отрыву электронов от определенных метаболитов с помощью специфических ферментов (дегидрогеназ) и акцептированию их другими молекулами, образующимися из сбраживаемого субстрата, т. е. в процессе брожения происходит окисление анаэробного типа.
Собственно энергетической стороной процессов брожения является их окислительная часть, поскольку реакции, ведущие к выделению энергии, - это реакции окисления. Существует несколько исключений из этого правила: некоторые анаэробы часть энергии при сбраживании субстрата получают также в результате его расщепления. Примитивность процессов брожения заключается в том, что из субстрата в результате его анаэробного преобразования извлекается лишь незначительная доля той химической энергии, которая в нем содержится. Продукты, образующиеся в процессе брожения, все еще содержат в себе значительное количество энергии, заключавшейся в исходном субстрате. Чтобы четче представить разницу в энергетическом выходе процессов брожения и дыхания, приведем данные по изменению уровней стандартной свободной энергии для процессов гомоферментативного молочнокислого брожения и дыхания при одинаковом исходном энергетическом субстрате (глюкоза):
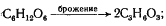
| 
|
| DG0' = –196,65 кДж/моль; | DG0' = –2870,22 кДж/моль. |
В процессе гомоферментативного молочнокислого брожения синтезируются 2 молекулы АТФ на 1 молекулу сброженной глюкозы; в процессе дыхания при полном окислении молекулы глюкозы образуется 38 молекул АТФ. В обоих случаях эффективность запасания выделяющейся энергии в макроэргических связях АТФ приблизительно одинакова.
При брожении некоторые реакции на пути анаэробного преобразования субстрата связаны с наиболее примитивным типом фосфорилирования - субстратным фосфорилированием. К синтезу АТФ по механизму субстратного фосфорилирования ведут катаболические реакции, которые в зависимости от своей химической природы могут быть разделены на два типа. Большинство относится к окислительно-восстановительным реакциям. Богатые энергией соединения возникают в процессе брожения на этапах анаэробного окисления. Например, окисление фосфоглицеринового альдегида (ФГА), катализируемое ФГА-дегидрогеназой, приводит к образованию богатого энергией метаболита - 1,3-дифосфоглицериновой кислоты (1, 3-ФГК). Анаэробное окисление пировиноградной или a-кетоглутаровой кислот приводит к образованию высокоэнергети-ческих метаболитов - ацетил-КоА или сукцинил-КоА соответственно.
Второй тип реакций связан с расщеплением субстратов или промежуточных продуктов, образующихся из них. Катализируются эти реакции ферментами, относящимися к классу лиаз. Например, у гетероферментативных молочнокислых бактерий высокоэнергетический ацетилфосфат образуется из ксилулозо-5-фосфата в реакции, катализируемой фосфокетолазой:
ксилулозо-5-фосфат + ФН → ФГА+ ацетилфосфат + H2O.
К реакциям подобного типа относится также расщепление цитруллина, приводящее к синтезу карбамоилфосфата — соединения с макроэргической фосфатной связью:
цитруллин + ФН → карбамоилфосфат + орнитин.
Богатые энергией соединения, образующиеся в реакциях рассмотренных выше типов, представляют в большинстве случаев ангидриды фосфорной кислоты или тиоэфиры органических кислот. Последние используются для синтеза АТФ через ферментативную стадию образования соответствующих ацилфосфатов:
ацил-КоА + ФН → ацилфосфат + КоА-SH.
Из других высокоэнергетических соединений важное место в энергетике процессов брожения принадлежит фосфоенолпировиноградной кислоте (ФЕП). Эти соединения характеризуются тем, что свободная энергия, освобождающаяся при их гидролизе, находится в области значений от -35 до -88 кДж/моль и с помощью соответствующих ферментов может быть перенесена на молекулы АДФ.
Несмотря на большое число углеродных субстратов, доступных для сбраживания, количество реакций, приводящих непосредственно к синтезу АТФ при брожениях, сравнительно невелико. Наиболее распространены следующие из них:
1) ацетилфосфат + АДФ → ацетат + АТФ;
2) 1,3-фосфоглицериновая кислота + АДФ → 3-фосфоглицеринновая кислота + АТФ;
3) фосфоенолпировиноградная кислота + АДФ → пировиноградная кислота + АТФ.
Другие реакции субстратного фосфорилирования ограничены какими-либо специфическими видами брожения. Например, сбраживание некоторых пиримидинов и аргинина, осуществляемое отдельными видами бактерий из рода Streptococcus, приводит к образованию карбамоилфосфата, фосфатная группа которого переносится на АДФ в реакции, катализируемой карбаматкиназой:
карбамоилфосфат + АДФ ® карбамат + АТФ.
Проблема акцептора электронов
Основная проблема всех процессов брожения — проблема акцептора электронов. В конечном итоге степень окисления и сопряженное с этим количество свободной энергии, а также характер образующихся продуктов определяются природой конечных акцепторов электронов. При брожениях конечными акцепторами электронов служат в основном органические соединения: метаболиты, образующиеся из исходных субстратов (пировиноградная кислота, ацетальдегид), или вещества, имеющиеся в среде культивирования (некоторые аминокислоты и другие органические соединения, способные восстанавливаться). В ряде брожений акцепторами электронов служат молекулы CO2, а также ионы водорода (H2). Кроме того, в отдельных случаях дополнительными акцепторами электронов могут быть некоторые достаточно окисленные неорганические соединения, такие как нитрат, молекулярная сера. Если конечным акцептором электронов является ацетальдегид, образуется этанол, если пируват — молочная кислота. Акцептирование электронов молекулами CO2 приводит у разных видов к возникновению формиата или ацетата, если же эту функцию выполняют ионы водорода, образуется молекулярный водород (H2).
Восстановленные соединения, акцептировавшие электроны, выделяются из клеток эубактерий в окружающую среду и накапливаются в ней в значительных количествах. Из-за низкого энергетического выхода процессов брожения для обеспечения энергией всех функций и биосинтетических процессов клетке приходится перерабатывать огромные количества субстратов.
Итак, брожение - это способ получения энергии, при котором АТФ образуется в процессе анаэробного окисления органических субстратов в реакциях субстратного фосфорилирования.
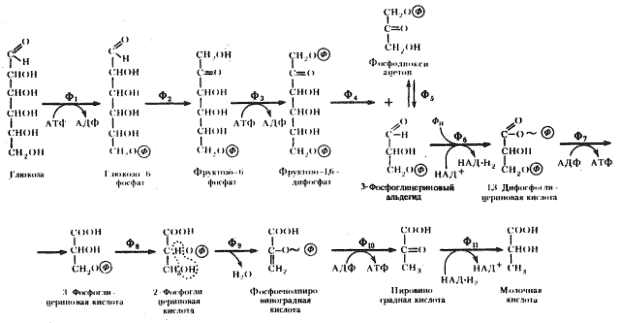
Гомоферментативное молочнокислое брожение. Последовательность биохимических реакций, лежащих в основе гомоферментативного молочнокислого брожения, получила название гликолитического пути (гликолиза).
|
Основными энергетическими ресурсами для эубактерий, осуществляющих гомоферментативное молочнокислое брожение, служат моносахара (в первую очередь, глюкоза) и дисахара (мальтоза, лактоза). В процессе подготовки к энергетическим преобразованиям дисахара ферментативным путем расщепляются до моносахаров. Различные моносахара, прежде чем подвергнуться преобразованиям, должны превратиться в глюкозо-6-фосфат. Момент унификации, т.е. превращения различных субстратов в один, исходный для дальнейшего его метаболизирования по данному пути, очень важен. От того, что служит исходным энергетическим ресурсом, зависит общий энергетический баланс процесса.
Собственно гликолиз - это определенная последовательность ферментативных реакций от углевода до пировиноградной кислоты, поэтому, строго говоря, «гликолиз» не является синонимом «гомоферментативного молочнокислого брожения», но 10 из 11 реакций у этих процессов идентичны.
Гомоферментативное молочнокислое брожение представляет собой энергетическую сторону образа жизни группы гомоферментативных молочнокислых бактерий. В процессе гомоферментативного молочнокислого брожения имеют место 3 типа химических превращений:
- перестройка углеродного скелета исходного субстрата;
- окислительно-восстановительные превращения;
- образование АТФ.
Энергетический выход процесса таков: образование 2 молекул АТФ на молекулу глюкозы. Энергетическая эффективность процесса, т. е. эффективность запасания выделяемой свободной энергии в молекулах АТФ, составляет примерно 40%. Энергия запасается только в реакциях субстратного фосфорилирования. Как можно видеть из суммирования энергетических характеристик процесса, низкий энергетический выход сочетается в нем с высокой энергетической эффективностью, а в основе всего лежат простые механизмы получения энергии.
Возникнув как первый, далекий от совершенства энергетический процесс, гомоферментативное молочнокислое брожение не было потом отброшено в процессе эволюции. Наоборот, оно закрепилось и существует сейчас в виде гликолиза у подавляющего большинства прокариот, дрожжей, грибов, а также у высших животных и растений, но только как первый этап более совершенного энергетического процесса, сформировавшегося в результате последующего развития способов получения энергии живыми организмами. Чем объясняется такая судьба гомоферментативного молочнокислого брожения? Вероятно, оказалось выгодным использовать его в качестве первого подготовительного этапа по следующим причинам: 1) высокая энергетическая эффективность (не путать с энергетическим выходом процесса!); 2) простота механизмов получения энергии; 3) перестройка исходного субстрата в форму, метаболически удобную для последующих превращений.
Гомоферментативное молочнокислое брожение, в основе которого лежит гликолитический путь разложения глюкозы, является единственным способом получения энергии для группы эубактерий, которые при сбраживании углеводов превращают в молочную кислоту от 85 до 90% сахара среды. Бактерии, входящие в данную группу, морфологически различны. Это кокки, относящиеся к родам Streptococcus и Pediococcus, а также длинные или короткие палочки из рода Lactobacillus. Все бактерии этой группы положительно окрашиваются по Граму, не образуют спор, неподвижны.
Молочнокислые бактерии распространены там, где они могут обеспечить свои высокие потребности в питательных веществах и где имеются большие количества углеводов, переработка которых дает им необходимую для роста энергию. Их много в молоке и молочных продуктах, на поверхности растений и в местах разложения растительных остатков; обнаружены они в пищеварительном тракте и на слизистых оболочках животных и человека.
Спиртовое брожение. Выше мы разобрали наиболее простой способ решения донор-акцепторной проблемы, который реализуется в виде молочнокислого брожения у группы гомоферментативных молочнокислых бактерий. Дальнейшие поиски на путях эволюции привели к формированию других метаболических возможностей для решения этой проблемы. Одна из них заключается в том, что из пировиноградной кислоты в результате ее окислительного декарбоксилирования образуется, ацетальдегид, который становится конечным акцептором водорода. В итоге из 1 молекулы гексозы образуются 2 молекулы этилового спирта и 2 молекулы углекислоты. Процесс получил название спиртового брожения. Спиртовое брожение распространено среди прокариотных (различные облигатно и факультативно анаэробные эубактерии) и эукариотных (дрожжи) форм. В анаэробных условиях у высших растений также отмечено накопление этилового спирта.
Процесс спиртового брожения, осуществляемый дрожжами, до последней реакции идет по тому же пути, что и описанный выше процесс молочнокислого брожения, но последняя реакция заменена двумя другими ферментативными реакциями. Сначала пируват с помощью пируватдекарбоксилазы, ключевого фермента спиртового брожения, декарбоксилируется до ацетальдегида и CO2:
CH3-CO-COOH → CH3-COH + CO2 .
Особенность реакции заключается в ее полной необратимости. Образовавшийся ацетальдегид восстанавливается до этанола с участием НАД+-зависимой алкогольдегидрогеназы:
CH3-COH + НАД-H2 → CH3-CH2OH + НАД+
Донором водорода служат 3-ФГА (как и в случае молочнокислого брожения).
Процесс спиртового брожения суммарно можно выразить следующим уравнением:
C6H12O6 + 2ФН + 2АДФ → 2CH3-CH2OH + 2CO2 + 2АТФ +2H2O.
Как видно из уравнения, с точки зрения энергетического выхода оба процесса (гомоферментативное молочнокислое и спиртовое брожение) одинаковы. В обоих случаях сбраживание 1 молекулы глюкозы приводит к образованию 2 молекул АТФ. Процессы различаются природой конечных акцепторов электронов. Кроме того, если при гомоферментативном молочнокислом брожении образовавшаяся молочная кислота в целом по степени окисленности-восстановленности не отличается от молекулы гексозы (имеет место лишь внутримолекулярное перераспределение окисленности и восстановленности отдельных углеродных атомов, входящих в ее молекулу), то в случае спиртового брожения происходит межмолекулярное размежевание на восстановленные (этиловый спирт) и окисленные (CO2) молекулы.
Накопление этилового спирта в среде в анаэробных условиях наблюдается у разных групп эубактерий и группы эукариотных микроорганизмов - дрожжей. Способность осуществлять в анаэробных условиях спиртовое брожение по пути, описанному в предыдущем разделе, присуща некоторым эубактериям, принадлежащим к разным таксономическим группам, например Sarcina ventriculi, Erwinia amylovora.
S. ventriculi относится к группе грамположительных анаэробных кокков. Клетки неподвижные; делятся в трех плоскостях, поэтому в культуре часто образуют пакеты, состоящие из 64 и более клеток. Веществом, связывающим клетки в пакетах, служит целлюлоза. Описана способность образовывать эндоспоры. Аэротолерантный анаэроб. Единственный способ получения энергии - сбраживание сахаров. Потребность в питательных веществах довольно высока (многочисленные аминокислоты и ряд витаминов). E. amylovora относится к группе энтеробактерий. Это грамотрицательные подвижные палочки. Особенностью вида является его патогенность для растений. Факультативный анаэроб. В аэробных условиях получает энергию в процессе дыхания.
Пропионовокислое брожение. Из рассмотренных двух типов брожения видно, что ключевым соединением в обоих процессах является пируват, поскольку в конечном итоге специфика брожения определяется дальнейшей судьбой пирувата. Основная задача последующих реакций - регенерирование молекулы НАД+ и возвращение ее в клеточный метаболизм. Прямое восстановление пирувата с помощью НАД-H2 до молочной кислоты реализуется в молочнокислом брожении. Другая возможность регенерирования НАД+ - «сбрасывание» водорода с НАД-H2 на фрагменты, образуемые при метаболизме пирувата, — имеет место в спиртовом брожении, осуществляемом дрожжами и некоторыми видами бактерий. Третья возможность связана с синтетическим процессом - усложнением молекулы пирувата, в результате которого создается более окисленная молекула акцептора, способная принять больше электронов с восстановленных переносчиков. Это происходит при присоединении к молекуле пирувата CO2, приводящем к формированию четырехуглеродного скелета. Процесс получил название гетеротрофной ассимиляции углекислоты.
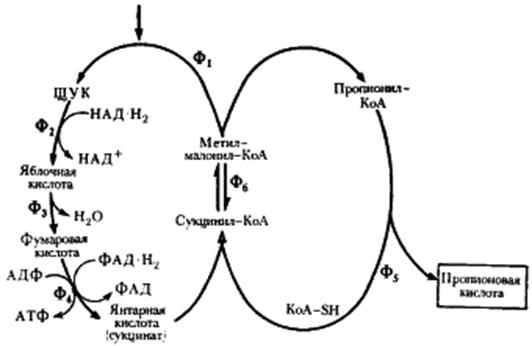
|
| Превращение пировиноградной кислоты в пропионовую при пропионовокислом брожении |
В пропионовокислом брожении мы имеем дело с реализацией третьей возможности превращения пирувата - его карбоксилированием, приводящим к возникновению нового акцептора водорода - ЩУК.
Энергетическая эффективность пропионовокислого брожения связана с выработкой пропионовыми бактериями новых метаболических способностей: реакций транскарбоксилирования и перегруппировки, участия в процессе КоА-производных. Образование дикарбоновой кислоты из пировиноградной с использованием механизма транскарбоксилирования вместо прямого карбоксилирования пирувата позволяет избежать дополнительных энергетических затрат на этом этапе брожения. Все это вместе взятое позволяет рассматривать пропионовокислое брожение как более совершенный из рассмотренных до сих пор способов получения энергии в анаэробных условиях.
Большинство пропионовокислых бактерий - аэротолерантные анаэробы, получающие энергию в процессе брожения, основным продуктом которого является пропионовая кислота. Аэротолерантность их обусловлена наличием полностью сформированной ферментной системы защиты от токсических форм кислорода (супероксидный анион, перекись водорода). У пропионовокислых бактерий обнаружены супероксиддисмутазная, каталазная и пероксидазная активности. Внутри группы отношение к O2 различно. Некоторые виды могут расти в аэробных условиях.
Местообитание пропионовых бактерий - кишечный тракт жвачных животных, молоко, твердые сыры, в приготовлении которых они принимают участие. После молочнокислого брожения, когда лактоза превращена в молочную кислоту, начинают размножаться пропионовые бактерии, сбраживающие молочную кислоту с образованием уксусной и пропионовой кислот. Эти кислоты придают сырам специфический острый вкус. Пропионовые бактерии используют в микробиологической промышленности в качестве продуцентов витамина B12.
Маслянокислое брожение. Следующий вариант решения донор-акцепторной проблемы на базе гликолитически образованного пирувата представляет собой маслянокислое брожение. Новое в маслянокислом брожении - возникновение реакций конденсации типа С2 + С2 → С4, в результате чего образуется С4-акцепторная кислота. Судьба этой кислоты различна и определяется необходимостью акцептирования водорода с НАД-H2, освобождающегося в процессе брожения, а это в свою очередь тесно связано с оттоком водорода на конструктивные процессы. В качестве конечных C4-продуктов в процессе брожения возникают соединения различной степени восстановленности. Характерным C4-продуктом брожения является масляная кислота. Осуществляют такой тип брожения многие бактерии, относящиеся к роду Clostridium.
Типичными представителями клостридиев, осуществляющих маслянокислое брожение, являются C. butyricum и C. pasteurianum. Они сбраживают сахара с образованием масляной и уксусной кислот, CO2 и H2. Превращение глюкозы до пирувата осуществляется по гликолитическому пути. Следующая реакция - разложение пирувата до ацетил-КоА и CO2, сопровождающееся образованием восстановленного ферредоксина (Фд). Реакция катализируется ферментом пируват: ферредоксиноксидоредуктазой и является ключевой в маслянокислом брожении. Особенности реакции - участие в ней белков, содержащих негемовое железо и кислотолабильную серу (FeS-белки).
Путь, ведущий к синтезу масляной кислоты, начинается с реакции конденсации двух молекул ацетил-КоА. Образовавшийся ацетоацетил-КоА восстанавливается в β-оксибутирил-КоА. Источником электронов в этой реакции и дальше на пути синтеза масляной кислоты служат молекулы НАД H2, образующиеся при окислении 3-ФГА в 1,3-ФГК.
Дальнейшее превращение заключается в отщеплении от молекулы (β-оксибутирил-КоА молекулы воды, что приводит к образованию соединения с двойной углеродной связью. Кротонил-КоА ферментативно восстанавливается в бутирил-КоА. Масляная кислота образуется в реакции переноса кофермента А с молекулы бутирил-КоА на ацетат. Эта реакция более «выгодна» для клетки, так как не приводит к потере энергии (в отличие от реакции простого гидролиза). Образующийся в реакции ацетил-КоА возвращается в метаболический поток и может быть использован для синтеза АТФ (реакция 7) или же вновь участвовать в последовательности реакций, ведущих к синтезу масляной кислоты (реакции 2 - 6).
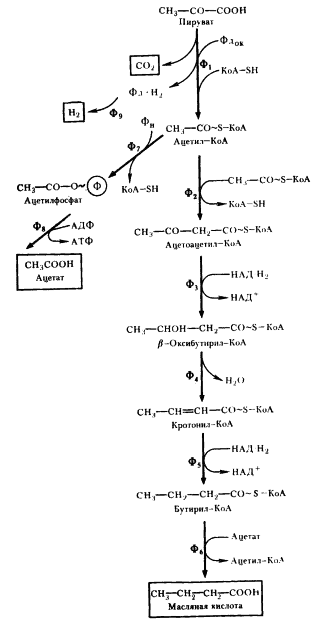
|
Разобранный выше путь, завершающийся синтезом масляной кислоты, не связан с получением клеткой энергии, поскольку ни на одном из этапов не происходит образования АТФ. Единственное назначение метаболических превращений ацетил-КоА по этому пути - акцептирование электронов, переносимых на НАД+ в процессе гликолитического метаболизирования глюкозы: две молекулы НАД-H2 образуются на этапе гликолиза, и на двух этапах превращений ацетил-КоА до масляной кислоты происходит потребление водорода с НАД-H2.
Расчеты показали, что в целом на 1 моль сбраживаемой глюкозы в маслянокислом брожении образуется 3,3 моля АТФ. Это наиболее высокий энергетический выход брожения, т.е. получения энергии за счет субстратного фосфорилирования, из всех рассмотренных выше типов брожений.
В природе есть много мест с полным или почти полным отсутствием молекулярного кислорода. Это глубокие слои воды, почвы, илы морей и континентальных водоемов. Особую экологическую нишу для развития анаэробов представляют рубец и кишечник животных и человека. Облигатно анаэробный способ существования широко распространен среди эубактерий. Систематическое изучение анаэробных эубактерий, предпринятое в последние десятилетия, обнаружило неоднородность входящих в эту группу организмов, способных получать энергию в процессах брожения, фотосинтеза и анаэробного дыхания.
Только небольшая часть облигатно анаэробных эубактерий может быть отнесена к первичным анаэробам, т. е. возникшим в докислородную эпоху и сохранившим до настоящего времени основные черты метаболизма того периода в результате обитания в анаэробных экологических нишах: получение энергии в процессе брожения, отсутствие электронтранспортных цепей, слабо развитые биосинтетические способности.
Большинство существующих облигатных анаэробов среди эубактерий имеют вторичное происхождение как следствие повторной адаптации к анаэробным условиям, сопровождающейся, как правило, изменениями деградационного характера: потерей способности взаимодействовать с O2, утратой некоторых компонентов переноса электронов, большей зависимостью от готовых органических соединений среды обитания и т. д. Примером могут служить строго анаэробные эубактерии, составляющие основную микрофлору рубца и пищеварительного тракта животных и человека. Это в большинстве грамотрицательные кокки или палочки, способные сбраживать сахара и/или аминокислоты. У многих из них обнаружены цитохромы b и a и показана способность синтезировать АТФ по механизму мембранзависимого фосфорилирования.
В представленном в этой главе материале проанализированы энергетические процессы, сформированные на первом этапе эволюции жизни на Земле. То, что брожение - наиболее примитивный способ получения энергии организмами, в настоящее время никем не ставится под сомнение. Гораздо сложнее оценить, какой путь в процессе эволюции пройден теми или иными организмами. Очевидно, что при имеющихся возможностях обмена генетической информацией в мире прокариот сохранение их в первоначальном виде маловероятно. Описание представленных в этой главе нескольких групп анаэробных эубактерий, в первую очередь, пропионовокислых бактерий и клостридиев, служит иллюстрацией этого.
Дата добавления: 2015-02-03; просмотров: 6775;