Генетический аппарат и репликация хромосомы
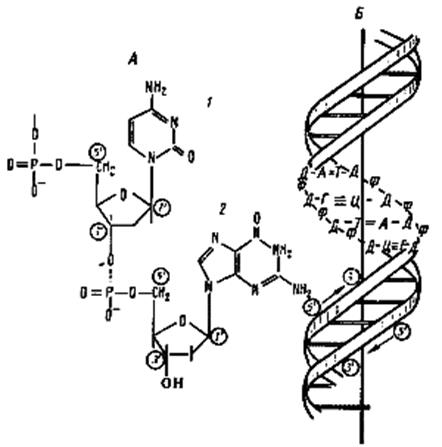
|
| Строение ДНК: А - фрагмент нити ДНК, образованной чередующимися остатками дезоксирибозы и фосфорной кислоты. К первому углеродному атому дезоксирибозы присоединено азотистое основание: 1 - цитозин; 2 - гуанин; Б - двойная спираль ДНК: Д - дезоксирибоза; Ф - фосфат; А - аденин; Т-— тимин; Г - гуанин; Ц - цитозин |
Строение генетического аппарата прокариот долгое время было предметом жарких дискуссий, суть которых сводилась к тому, есть у них такое же ядро, как у эукариот, или нет. Установлено, что генетический материал прокариотных организмов, как и эукариотных, представлен ДНК, но имеются существенные различия в его структурной организации. У прокариот ДНК представляет собой более или менее компактное образование, занимающее определенную область в цитоплазме и не отделенное от нее мембраной, как это имеет место у эукариот. Чтобы подчеркнуть структурные различия в генетическом аппарате прокариотных и эукариотных клеток, предложено у первых его называть нуклеоидом в отличие от ядра у вторых.
При электронно-микроскопическом наблюдении видно, что нуклеоид прокариот, несмотря на отсутствие ядерной мембраны, довольно четко отграничен от цитоплазмы, занимает в ней, как правило, центральную область и заполнен нитями ДНК диаметром около 2 нм. Не исключено, что на выявляемую в электронном микроскопе организацию прокариотной хромосомы большое влияние оказывают условия фиксации препарата. По имеющимся наблюдениям, в живой клетке нуклеоид занимает больше места в цитоплазме.
Вся генетическая информация прокариот содержится в одной молекуле ДНК, имеющей форму ковалентно замкнутого кольца и получившей название бактериальной хромосомы (в прокариотной клетке ДНК может находиться и вне бактериальной хромосомы - в плазмидах, но последние не являются обязательными клеточными компонентами.). Длина молекулы в развернутом виде может составлять более 1 мм, т.е. почти в 1000 раз превышать длину бактериальной клетки. Длительное время считали, что в распределении нитей ДНК бактериальной хромосомы не прослеживается никакой закономерности. Однако если исходить из того, что молекула ДНК образует беспорядочный клубок, трудно объяснить процесс репликации и последующее распределение образовавшихся хромосом по дочерним клеткам. Специальные исследования показали, что хромосомы прокариот представляют собой высокоупорядоченную структуру, имеющую константу седиментации 1300-2000S для свободной и 3200-7000S для связанной с мембраной формы. В том и другом случае часть ДНК в этой структуре представлена системой из 20-100 независимо суперспирализованных петель. В обеспечении суперспирализованной организации хромосом участвуют молекулы РНК.
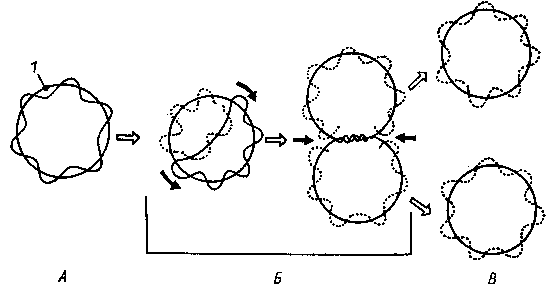
Репликация кольцевой бактериальной хромосомы в двух направлениях. А - родительская молекула ДНК; Б - промежуточные репликативные формы; В - дочерние молекулы ДНК после завершения процесса репликации и расхождения: 1-— точка начала репликации; черными стрелками показано направление репликации.
Хромосомы большинства прокариот имеют молекулярную массу в пределах 1-3 109 Да. В группе микоплазм генетический материал представлен молекулами, имеющими наименьшее для клеточных организмов количество ДНК (0,4-0,8 109), а наибольшее содержание ДНК обнаружено у нитчатых цианобактерий (8,5 109). Хотя каждая прокариотная клетка содержит 1 хромосому, часто в экспоненциально растущей культуре количество ДНК на клетку может достигать массы 3, 4, 8 и более хромосом. Нередко в клетках при действии на них определенных факторов (температуры, pH среды, ионизирующего излучения, солей тяжелых металлов, некоторых антибиотиков и др.) происходит образование множества копий хромосомы. При устранении воздействия этих факторов, а также после перехода в стационарную фазу в клетках, как правило, обнаруживается по одной копии хромосомы.
ДНК прокариот построена так же, как и эукариот. Молекула ДНК несет множество отрицательных зарядов, поскольку каждый фосфатный остаток содержит ионизированную гидроксильную группу. У эукариот отрицательные заряды нейтрализуются образованием комплекса ДНК с основными белками - гистонами. В клетках подавляющего большинства прокариот не обнаружено гистонов, поэтому нейтрализация зарядов осуществляется взаимодействием ДНК с полиаминами (спермином и спермидином), а также с ионами Mg2+. В последнее время у некоторых архебактерий и цианобактерий обнаружены гистоны и гистоноподобные белки, связанные с ДНК. Содержание пар оснований А+Т и Г+Ц в молекуле ДНК является постоянным для данного вида организма и служит важным диагностическим признаком. У прокариот молярная доля ГЦ в ДНК колеблется в очень широких пределах: от 23 до 75%.
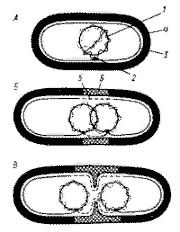
|
| Механизм распределения бактериальных хромосом: А - бактериальная клетка содержит частично реплицированную хромосому, прикрепленную к мембране в точке (или точках) репликации; Б- репликация хромосомы завершена. В бактериальной клетке две дочерние хромосомы, каждая из которых прикреплена к ЦПМ. Показан синтез клеточной стенки и ЦПМ; В- продолжающийся синтез мембраны и клеточной стенки приводит к разделению дочерних хромосом. Показано начало деления клетки путем образования поперечной перегородки: 1 - ДНК; 2 - прикрепление хромосомы к ЦПМ: 3 - ЦПМ; 4 - клеточная стенка: 5- синтезированный участок ЦПМ; 6 - новый материал клеточной стенки |
Деление молекулы ДНК (репликация) происходит по полуконсервативному механизму и в норме всегда предшествует делению клетки. С помощью электронного микроскопа установлено, что репликация ДНК начинается в точке прикрепления кольцевой хромосомы к ЦПМ, где локализован ферментативный аппарат, ответственный за репликацию. Часто можно обнаружить, что контакт ДНК с ЦПМ осуществляется посредством мезосом. Репликация, начавшаяся в точке прикрепления, идет затем в двух противоположных направлениях, образуя характерные для кольцевой хромосомы промежуточные структуры. Возникающие дочерние хромосомы остаются прикрепленными к мембране. Репликация молекул ДНК происходит параллельно с синтезом мембраны в области контакта ДНК с ЦПМ. Это приводит к разделению (сегрегации) дочерних молекул ДНК и оформлению обособленных хромосом
Модель строения бактериальной хромосомы должна объяснять также прохождение в клетке процессов транскрипции и трансляции. Согласно существующим представлениям суперспирализованные петли соответствуют неактивным в данное время участкам ДНК и находятся в центре нуклеоида. По его периферии располагаются деспирализованные участки, на которых происходит синтез информационной РНК (иРНК), при этом, поскольку у бактерий процессы транскрипции и трансляции идут одновременно, одна и та же молекула иРНК может быть одновременно связана с ДНК и рибосомами).
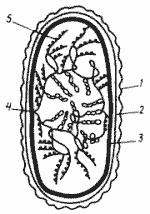
|
| Модель организации нуклеоида Е. coli - наружная мембрана клеточной стенки; 2 - пептидогликановый слой; 3 - ЦПМ; 4 - точка прикрепления бактериальной хромосомы к ЦПМ; 5 - рибосомы, «сидящие» на иРНК. |
Рост и способы размножения. Под ростом прокариотной клетки понимают согласованное увеличение количества всех химических компонентов, из которых она построена. Рост является результатом множества скоординированных биосинтетических процессов, находящихся под строгим регуляторным контролем, и приводит к увеличению массы (а следовательно, и размеров) клетки. Но рост клетки не беспределен. После достижения определенных (критических) размеров клетка подвергается делению.
Для подавляющего большинства прокариот характерно равновеликое бинарное поперечное деление, приводящее к образованию двух одинаковых дочерних клеток. При таком способе деления имеет место симметрия в отношении продольной и поперечной оси. У большинства грамположительных эубактерий и нитчатых цианобактерий деление происходит путем синтеза поперечной перегородки, идущего от периферии к центру. Так у Bacillus subtilis в середине клетки сначала имеет место кольцевое впячивание ЦПМ, сопровождающееся формированием мезосом разного внешнего вида. Они образуются в месте закладки поперечной перегородки, и предполагается их активное участие в процессах синтеза пептидогликана и других компонентов клеточной стенки. Поперечная перегородка формируется из ЦПМ и пептидогликанового слоя, ее наружные слои синтезируются позднее. Клетки большинства грамотрицательных эубактерий делятся путем перетяжки. У Е. coli на месте деления обнаруживается постепенно увеличивающееся и направленное внутрь искривление ЦПМ и клеточной стенки. Синтез новой клеточной стенки может происходить в нескольких местах или только в зоне формирования поперечной перегородки.
Вариантом бинарного деления является почкование, которое можно рассматривать как неравновеликое бинарное деление. При почковании на одном из полюсов материнской клетки образуется маленький вырост (почка), увеличивающийся в процессе роста. Постепенно почка достигает размеров материнской клетки, после чего отделяется от последней. Клеточная стенка почки полностью синтезируется заново. В процессе почкования симметрия наблюдается в отношении только продольной оси. При равновеликом бинарном делении материнская клетка, делясь, дает начало двум дочерним клеткам и сама, таким образом, исчезает. При почковании материнская клетка дает начало дочерней клетке, и между ними можно в большинстве случаев обнаружить морфологические и физиологические различия: есть старая материнская клетка и новая дочерняя. В этом случае можно наблюдать процесс старения. Так, для некоторых штаммов Rhodomicrobium показано, что материнская клетка способна отпочковывать не более 4 дочерних клеток. Дочерние клетки лучше приспосабливаются к меняющимся условиям. Почкование обнаружено в разных группах прокариот: среди фото- и хемотрофов, осуществляющих авто- и гетеротрофный конструктивный метаболизм. Вероятно, оно в процессе эволюции возникало несколько раз.
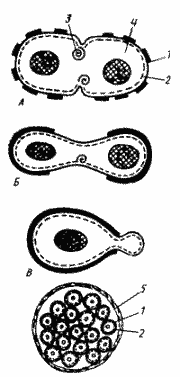
|
| Способы деления и синтез клеточной стенки у прокариот: А - деление путем образования поперечной перегородки; Б - деление путем перетяжки; В - почкование; Г - множественное деление: 1 - клеточная стенка (толстой линией обозначена клеточная стенка материнской клетки, тонкой — заново синтезированная); 2 - ЦПМ; 3 — мембранная структура; 4 - цитоплазма, в центре которой расположен нуклеоид; 5 - дополнительный фибриллярный слой клеточной стенки |
Бинарное деление может происходить в одной или нескольких плоскостях. В первом случае, если после деления клетки не расходятся, это приводит к образованию цепочек палочковидных или сферических клеток, во втором - к клеточным скоплениям разной формы). Расхождение образовавшихся дочерних клеток происходит в результате лизиса среднего слоя клеточной стенки.
Для одной группы одноклеточных цианобактерий описано размножение путем множественного деления. Оно начинается с предварительной репликации хромосомы и увеличения размеров вегетативной клетки, которая затем претерпевает ряд быстрых последовательных бинарных делений, происходящих внутри дополнительного фибриллярного слоя материнской клеточной стенки. Это приводит к образованию мелких клеток, получивших название баеоцитов (баеоцит - по-гречески маленькая клетка), число которых у разных видов колеблется от 4 до 1000. Освобождение баеоцитов происходит путем разрыва материнской клеточной стенки. Таким образом, в основе множественного деления лежит принцип равновеликого бинарного деления. Отличие заключается в том, что в этом случае после бинарного деления не происходит роста образовавшихся дочерних клеток, а они снова подвергаются делению.
Деление прокариотной клетки начинается, как правило, спустя некоторое время после завершения цикла репликации молекулы ДНК. Вероятно, репликация бактериальной хромосомы запускает какие-то процессы, ведущие к клеточному делению. Более детальное изучение у разных видов прокариот взаимосвязи между репликацией ДНК и делением клетки не привело к однозначным результатам. Получены данные о том, что сигналом к клеточному делению служит начало репликации ДНК, ее завершение или репликация определенного локуса бактериальной хромосомы. Таким образом, в норме существует вполне определенная временная связь между репликацией хромосомы и делением бактериальной клетки. Воздействия различными химическими веществами и физическими факторами, приводящие к подавлению репликации ДНК, останавливают и клеточное деление. Однако при некоторых условиях связь между обоими процессами может быть нарушена, и клетки способны делиться в отсутствие синтеза ДНК. Это удалось получить введением определенных мутаций в генетический аппарат бактериальной клетки.
Нарушить последовательность процессов репликации бактериальной хромосомы и клеточного деления также можно, выращивая бактерии при разной температуре. Культивирование Bacillus subtilis на богатой питательной среде при 37° приводит к интенсивному делению бактериальной хромосомы и росту клеток, в результате чего в культуре образуются нитевидные клетки, содержащие множество хромосомных копий с отсутствующими совсем или недосформированными (незамкнутыми) поперечными перегородками. При замедлении скорости роста наблюдается деление нитевидных клеток, приводящее к образованию бактериальных клеток нормальной длины.
Дата добавления: 2015-02-03; просмотров: 3540;