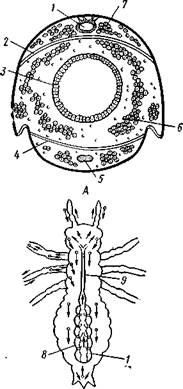
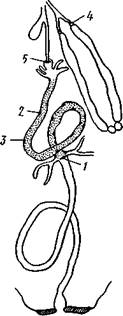
Аорта, 2 — сердце. 3 — остии, 4 — крыловидные мышцы
Важная физиологическая особенность дыхательной системы насекомых состоит в следующем. Обыкновенно кислород воспринимается животным в определенных участках его тела и оттуда разносится кровью по всему организму. У насекомых же воздухоносные трубочки пронизывают все тело и доставляют кислород прямо к местам его потребления, т. е. к тканям и клеткам, как бы заменяя собой кровеносные сосуды.
Кровеносная система в связи с отмеченной особенностью дыхательной системы развита у насекомых сравнительно слабо. В брюшке над кишечником залегает длинное трубковидное сердце (рис. 343). Задний конец его слепо замкнут, а полость поделена перегородками, несущими клапанные отверстия, на несколько камер (чаще 8). В стенках сердца залегают мышечные волокна, обеспечивающие его сокращение. Каждая камера снабжена парой боковых остий. На переднем конце сердце продолжается в мускулистую головную аорту, которая, достигнув мозга,
|
заканчивается отверстием, так что гемо-лимфа из нее поступает прямо в полость тела.
| Рис. 344. Кровообращение у насекомых. А — схема расположения сердца и диафрагм в теле насекомого (из Шванвича); Б -г- схема кровообращения у личинки стрекозы (по Кольбе):- / — сердце, 2 — верхняя диафрагма, 3—кишка, 4—нижняя диафрагма, 5 — брюшная нервная цепочка, 6 — жировое тело, 7 — перикардиальные клетки, 8—мышцы, лежащие под сердцем, 9 — аорта. Стрелки показывают направление тока гемо-лифмы |
Сердце окружено участком полости тела, или перикардиальным синусом, который отделен от остальной полости очень тонкой и во многих местах продырявленной перегородкой — верхней диафрагмой. С ней соединяется система парных крыловидных мышц, расположенных под сердцем. У многих насекомых часто имеется аналогичная перегородка и в нижней части тела под кишечником (рис. 344,Л). Совместная работа сердца и диафрагм обеспечивает циркуляцию гемо-лимфы по телу насекомого (рис. 344,5). Крыловидные мышцы, сокращаясь, опускают верхнюю диафрагму вниз и соответственно увеличивают объем перикар-диального синуса. Гемолимфа устремляется в его полость, а оттуда через остии в сердце. Волнообразное сокращение, пробегающее от заднего конца сердца к переднему, гонит гемолимфу в голову животного, где она и изливается в полость тела через отверстие аорты. Работа нижней диафрагмы создает ток гемолимфы к заднему концу тела. Число пульсаций сердца крайне варьирует не только у многих видов, но даже у одного и того же насекомого при разных его физиологических состояниях. Так, у бражника — Sphinx ligustri в покоящемся состоянии сердце сокращается 60—70, а во время полета— 140—150 раз в минуту.
Гемолимфа насекомых представляет бесцветную или желтоватую жидкость. В редких случаях, как, например, у личинок комаров Tendipes, растворенный гемоглобин придает ей ярко-красный цвет. Гемолимфа содержит фагоциты, а также

Рис. 345. Пяденица-обдирало Erannis defoliaria (1 — самец, 2 — самка, 3 — гусеница). Оранжевый обдирало Erannis aurantiaria (4 — самец, 5 — самка). Зимняя пяденица Operophtera brumata (6 — гусеница, 7 — самец, 8 — самка) (из Брема)
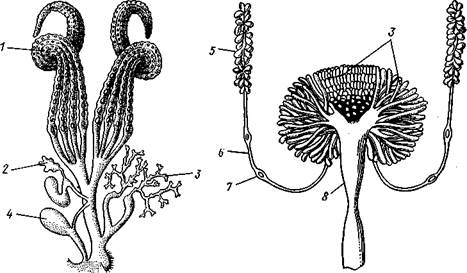
Рис. 346. Строение половой системы насекомых. А — женский половой аппарат
тутового шелкопряда Bombyx mori (по Жирару); Б — мужской половой аппарат
черного таракана Blatta orientalis (из Майаль и Денни):
Яичники, состоящие каждый из четырех яйцевых трубочек, 2 — семяприемник, 3 — придаточные железы, 4 — совокупительная сумка, соединяющаяся особым каналом с семяприемником; наружные отверстия совокупительной сумки и влагалища лежат рядом, 5—семенник, 6 — семяпровод, 7 — его раздвоение, 8 — семяизвергательяый канал
специальные кровяные клетки. Основная функция гемолимфы — снабжение тканей и органов питательными веществами. Кроме того, в нее поступают растворенные продукты обмена, которые переносятся к органам выделения. Дыхательная функция гемолимфы в связи с развитием трахейной системы невелика.
Половая система. Насекомые раздельнополы и нередко обладают резко выраженным половым диморфизмом. Он сказывается в меньшей величине самца (у многих бабочек) или в совершенно иной расцветке обоих полов (многие бабочки), или же в еще более существенных признаках. Так, например, у самцов появляются на теле различные придатки (рог жука-носорога) или же некоторые органы получают у самцов более сильное развитие, чем у самок (громадные жвалы жука-оленя, более длинные усики дровосеков). Иногда только самцы крылаты, тогда как у самок крылья редуцируются или вовсе исчезают (некоторые пяденицы, рис. 345; бабочки-мешочницы).
Половые железы парны. У самок (рис. 346, А) каждый яичник состоит большей частью из известного числа яйцевых трубок, сидящих, как пальцы на руке, на общем выводном канале яичника — яйцеводе. Тонкий слепой конец каждой яйцевой трубки состоит из скопления мелких однородных зачатковых клеток. Далее по направлению к яйцеводу яйцевая трубка постепенно расширяется и делится на ряд яйцевых камер. Каждая камера содержит крупную яйцевую клетку, окруженную слоем особого фолликулярного эпителия, клетки которого перед выходом яйца наружу выделяют вокруг него оболочку. Между соседними яйцами находится по группе питательных клеток, которые при росте яйца потребляются им. По мере роста яйцо продвигается в лицевой трубке по направлению к яйцеводу, а на смену выводимым яйцам на слепом конце трубки из зачатковых клеток идет формирование новых яиц и питательных клеток.
Оба яйцевода сливаются в непарное влагалище, открывающееся наружу на брюшной стороне брюшка под порошицей. Во влагалище открывается еще особый мешочек, семяприемник, где хранится семя после оплодотворения, а нередко, кроме того, мускулистая совокупительная сумка, служащая для введения в нее совокупительного органа самца. Семя переводится затем из сумки в семяприемник. У некоторых насекомых совокупление происходит всего один раз за несколько лет, и все это время живчики сохраняются живыми в семяприемнике самки, например у царицы пчел, жизнь которой длится 4—5 лет.
Нередко в области полового отверстия развиваются особые придатки — яйцеклады (с. 350), служащие для откладки яиц внутрь какого-нибудь более или менее твердого субстрата (почва, ткани растений и т. п.).
Мужская половая система (рис. 346, Б) состоит из пары простых или разделенных на несколько долек семенников. Два отходящих от них семяпровода сливаются в общий семяизвергательный канал; в последний очень часто открываются, кроме того, парные дополнительные железы. Семяизвергательный канал пронизывает совокупительный орган, образованный цилиндрическим выростом стенки тела, расположенным в небольшом впячивании, клоаке, куда открывается и порошица. У насекомых, не имеющих совокупительного органа, живчики склеиваются в сперматофоры, одетые сложной оболочкой, и в таком виде вводятся в половое отверстие самки.
Развитие.У насекомых развитие делится на два периода — эмбриональный и постэмбриональный.
Эмбриональное развитие. Яйцевые клетки насекомых имеют большей частью овальную форму и очень богаты желтком, который заполняет
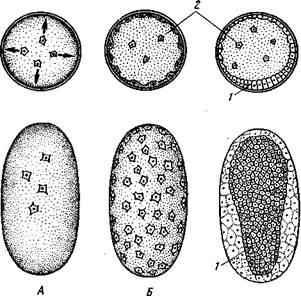
всю центральную часть яйца. Соответственно с этим дробление яйца частичное, поверхностное (рис. 347); в результате образуется сплошной слой одинаковых клеток, бластодерма, покрывающая центральный желток. Часть ядер при этом остается в желточной массе: в дальнейшем за их счет образуются особые желточные клетки (рис. 347). На будущей
|
| В |
Рис. 347. Дробление и образование зародышевой полоски у насекомых. А — начало дробления; Б — начало образования бластодермы; В — формирование зародышевой полоски: верхний ряд — поперечные разрезы через зародыши; нижний ряд — вид зародыша
с брюшной стороны (из Иоффа): / — зародышевая полоска, 2 — желточные клетки
брюшной стороне зародыша бластодерма утолщается. Этот участок называется зародышевой полоской; в дальнейшем за его счет формируется главная часть тела зародыша.
В области зародышевой полоски начинаются процессы дифференци-ровки. В результате размножения и ухода в глубь клеток бластодермы формируются две полоски клеток — мезодермальные полоски, а сама бластодерма становится эктодермой. Позднее мезодермальные полоски делятся на парные зачатки, дающие начало двойному ряду целомических мешочков. У высших насекомых (отр. Diptera) сегментация мезодер-мальных полосок подавляется и обособления самостоятельных целомов не происходит.
Вопрос о формировании энтодермы у насекомых еще не решен полностью. У многих форм зачатком энтодермы, по-видимому, являются желточные клетки. У Diptera и некоторых других насекомых на переднем и заднем концах зародышевой полоски обособляются две группы клеток, которые также рассматриваются как зачатки энтодермы.
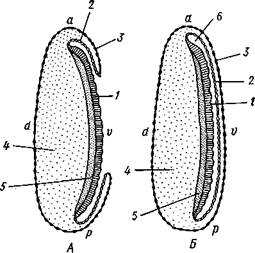
Еще в самом начале закладки зародышевых пластов начинается и крайне характерный для развития насекомых процесс формирования зародышевых оболочек (рис. 348).
В самом простом и широко распространенном случае по периферии
|
зародышевой полоски эктодерма образует сначала валик, а затем двухслойную складку, постепенно нарастающую со всех сторон на зародышевую полоску, которая вместе с тем несколько погружается в желток. Наконец, края складки смыкаются над зачатком зародыша, оставляя между собой и им занятую жидкостью амниотическую полость, отделенную от внешней среды двойной оболочкой. Наружный слой оболочки происходит в результате слияния наружных слоев смыкающейся над зародышем эктодермаль-ной складки и называется се-розой, внутренний — от внут-
реннего слоя той же складки, последний непосредственно ограничивает амниотическую полость и называется амнионом. Сероза и амнион защищают зародыш от повреждений.
| Рис. 348. Схема последовательных стадий (А, Б) образования зародышевых оболочек у насекомых (по Гейдеру): / — зародышевая полоска, 2 — амнион, 3 — сероза, 4 — желток, 5 — общий зачаток энтодермы и мезодермы, 6 — амниотическая полость. а — передний, р — задний полюс, d — спинная и v — брюшная стороны |
Между тем зародышевая полоска испытывает сегментацию, причем сегменты обособляются, как и у прочих Arthro-poda и кольчецов, в порядке спереди назад (рис. 349, А, Б). В головном отделе закладываются глазные и антен-нальные лопасти с соответствующими зачатками, интерка-лярный сегмент, впоследствии подвергающийся редукции, и
3 сегмента ротовых конечностей. Далее кзади дифференцируются 3 грудных, несущих зачатки конечностей, и 11 брюшных сегментов.
Что касается дифференцировки внутренних органов, то в главных чертах она заключается в следующем. Передняя и задняя кишки образуются при помощи двух эктодермальных впячиваний. Более сложен вопрос происхождения средней кишки. Для насекомых характерно удивительное разнообразие способов и источников образования последней. В одних случаях она формируется за счет желточных энтодермальных клеток, в других — в образовании средней кишки принимают участие эктодермальные элементы — специальные клетки, локализующиеся на слепо замкнутых концах зачатков передней и задней кишок. И, наконец, у ряда видов (партеногенетические самки тлей — см. с. 391, некоторые наездники — с. 385) средняя кишка формируется только за счет эктодермы благодаря сильному разрастанию зачатка передней кишки, который достигает заднего конца тела. Энтодермальные элементы при этом дегенерируют. Мальпигиевы сосуды возникают из эктодермы задней кишки. Нервная система закладывается в виде брюшного валика эктодермы, который затем погружается под покровы. Целомические мешки распадаются, образуя смешанную полость тела — миксоцель. Освобождающиеся при этом клеточные элементы дают начало мышцам, сердцу, жировому телу и некоторым другим органам. Первичные половые клетки закладываются очень рано, иногда еще в тот период, когда желток окружен однослойной бластодермой.
Зародыш растет, питательный желток постоянно потребляется, и зародышевые оболочки лопаются, заворачиваются на спинную сторону в виде складочек, которые подвергаются дегенерации. Зародыш готов к выходу из яйца, и этим эмбриональное развитие насекомого заканчивается.
Постэмбриональное развитие. У насекомых из яйца выходит молодое животное или личинка с полным числом сегментов. Исключение составляют только примитивные скрыточелюстные, относящиеся к отр. Protura, у которых молодь отличается от взрослых отсутствием трех задних сегментов брюшка. Последние формируются из задней зоны роста последовательно после первых линек. Таким образом, для насекомых характерно развитие без анаморфоза (с. 338) и лишь у протур встречается остаточный анаморфоз.
Постэмбриональное развитие насекомых характеризуется значительным разнообразием, которое во многом предопределяется биологическими особенностями представителей этой группы животных.
У наиболее примитивных форм (подкл. Entognatha) рост и развитие молодого организма не сопровождаются существенными изменениями строения. Молодь во всех главных чертах похожа на материнский организм, т. е. личиночная стадия и метаморфоз отсутствуют. По сущест"
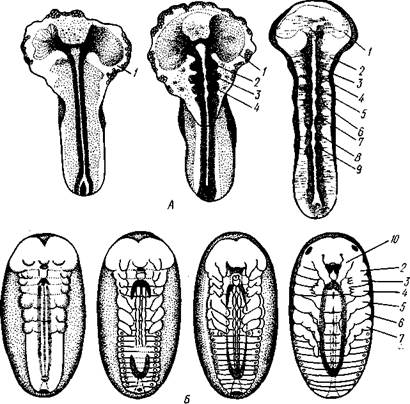
Рис. 349. Сегментация зародыша насекомого. А — три последовательные стадии
сегментации зародышевой полоски жука Melasoma (по Коршильту и Гейдеру);
6 — схема сегментации зародыша и закладки конечностей (из Иоффа):
/ — головная доля, 2—4 — зачатки челюстных сегментов, 5—7 — зачатки грудных сегментов, 8, 9 — зачатки первых двух брюшных сегментов, 10 — зачатки сяжков
ву, б этом случае мы имеем дело с прямым развитием. Крылатые насекомые, стоящие более высоко в эволюционном отношении, напротив, обладают метаморфозом, причем по характеру превращения они делятся на гемиметаболические — с неполным превращением и голометаболи-ческие — с полным метаморфозом. Неполный метаморфоз характерен
|
| 321 |
| «SD |
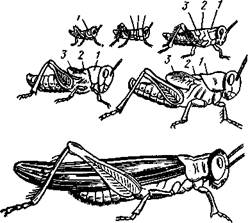
для более низко организованных групп крылатых насекомых: прямокрылые, тараканы, клопы, поденки, стрекозы. При неполном превращении (рис. 350) вылупляющаяся из яйца личинка по общему облику во многом уже похожа на взрослый организм, отличаясь от него главным образом зачаточным состоянием крыльев, недоразвитием вторич-нополовых признаков и нередко наличием некоторых провизорных, чисто личиночных органов: например, трахейные жабры у личинок поденок и др. Крылья имеются сначала лишь в виде едва заметных наружных зачат-
Рис. 350. Неполный метаморфоз у саранчи ков. Сглаживание разницы меж-
Locusta migratoria (из Холодковского): Ду личинкой и взрослым ОрГаНИЗ-
^а- сегменты груди MQM прОИСХОДИТ ПОСТеПСННО,
сопровождаясь рядом линек.
С каждой последующей линькой личинка все более становится похожей на взрослую стадию насекомых, или имаго (imago), зачатки крыльев увеличиваются и т. д.
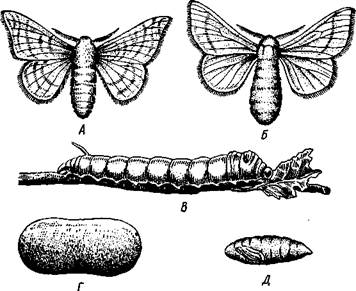
Рис. 351. Полный метаморфоз у тутового шелкопряда Bombyx то-
ri. А — самец; Б — самка; В — личинка (гусеница); Г — кокон;
Д — куколка, вынутая из кокона (по Лейнису)
У некоторых примитивных насекомых количество линек очень велико и может достигать 25—30 (поденки). У представителей других отрядов линек меньше (4—5).
У голометаболических насекомых развитие иное (рис. ^51). Личинка резко отличается от взрослого животного: она вовсе не имеет наружных зачатков крыльев, нередко имеет другое число конечностей и т. д. Личиночные признаки сохраняются в течение ряда линек, т. е. во время всего личиночного периода жизни. Личинки голометаболических насекомых чрезвычайно разнообразны. Личинка отличается от имаго наличием ряда примитивных признаков: большей гомономностыо расчленения, отсутствием крыльев, слабым развитием органов чувств (сложных глаз нет, а имеются лишь глазки, антенны очень малы и т. д.), а во многих случаях также устройством и количеством конечностей. Очень часто ротовые конечности в личиночном возрасте устроены по одному типу (грызущие у гусениц бабочек), у взрослых — по другому (сосущий ротовой аппарат у взрослых бабочек). Количество и строение ног также отличается у личинок разных систематических групп. У многих имеются только три пары конечностей на грудных сегментах (личинки жуков и др.). У гусениц бабочек и личинок некоторых перепончатокрылых (пилильщики, с. 397) на брюшных сегментах развивается еще несколько пар конечностей — ложные ножки; последние отличаются слабой расчлененностью и наличием присоски. У взрослых насекомых они не сохраняются. И наконец, встречаются фактически безногие личинки, у которых не только грудные, но и часть ротовых конечностей может редуцироваться, так что тело принимает вид малоподвижного и неясно расчлененного мешка. Это характерно для личинок, которые либо выкармливаются взрослыми особями (многие перепончатокрылые), либо обитают на субстрате, используемом как пища. Таковы, например, паразитические личинки и формы, поселяющиеся в различных разлагающихся органических веществах (многие двукрылые — отр. Diptera).
Как уже сказано, строение, характерное для личинки, сохраняется в течение всего личиночного периода: превращение совершается не постепенно, а необходимые для метаморфоза изменения отодвигаются ко времени последней личиночной линьки. В этот период совершаются такие коренные преобразования организма личинки, что продолжение обычных физиологических отправлений, особенно питания и движения, становится затруднительным, даже невозможным. Это сказывается в том, что личинка после последней линьки переходит в состояние покоя — становится куколкой. Куколка составляет наиболее характерную стадию го-лометаболического превращения. Чем полнее состояние покоя, тем лучше выражено полное превращение.
Различают три типа куколок (рис. 352). У свободных куколок зачатки крыльев и конечностей взрослого насекомого явственно видны и свободно выдаются над поверхностью тела. У покрытых куколок зачатки так тесно приложены и как бы приклеены к туловищу, что заметны лишь в виде неясных контуров. Куколки этих двух типов в определенной мере подвижны: куколки комаров, например, за счет изгибаний брюшка плавают в воде. Куколки многих жуков и бабочек также способны изгибать брюшко (иногда довольно энергично), но направленным движением не обладают. Третья категория — совершенно неподвижные куколки, встречаются у части мух; тело такой куколки бесформенного, бочонкообразно-го вида вследствие того, что остается окруженным отставшей, но не сброшенной кожицей от последней личиночной линьки.
В течение описываемой стадии покоя внутри куколки происходят важные изменения организации, подготавливающие переход в стадию
имаго. Так как и род движения (полет), и питание у имаго большей частью отличны от таковых личиночного периода, то и мышцы, и пищеварительный канал личинки должны подвергнуться полной перестройке. Большая часть органов и тканей тела личинки во время стадии куколки совершенно переформировывается. Органы личинки разрушаются, пре-
|
Рис. 353. Кишечник куколки мухи с имагинальными ^
дисками (по Ковалевскому):
/ — кольцевидный диск для задней кишки, 2, 3 — имагиналь-
Дата добавления: 2015-01-24; просмотров: 1987;