Основание конечности
ний и нижний листки пластинки сближаются, мягкие ткани частично дегенерируют, и крыло приобретает вид тонкой перепонки.
Движение крыльев у насекомых — результат работы сложного механизма и определяется, с однойстороны, особенностью сочленения крыла с туловищем, а с другой — действием особых крыловых мышц. В общих чертах основной механизм движения крыльев представляется следующим образом (рис. 319). Само крыло — это двуплечий рычаг с неодинаковой длиной плечей. С тергитом и боковой пластинкой крыло соединяется тонкими и гибкими мембранами. Чуть отступя от места этого соединения, крыло опирается на небольшой, имеющий вид столбика вырост боковой пластинки, который и является точкой опоры крылового рычага.
Расположенные в грудных сегментах мощные продольные и дорзо-вентральные мышцы могут опускать или приподнимать тергит. При опускании последний давит на короткое плечо крыла и влечет его за собой вниз. В результате длинное плечо, т. е. вся несущая плоскость крыла, движется вверх. Подъем тергита приводит к опусканию крыловой пластинки (рис. 319). Небольшие мышцы, прикрепленные непосредственно к крылу, способны поворачивать его вдоль продольной оси, при этом изменяется угол атаки. Во время полета свободный конец крыла движется по довольно сложной траектории (рис. 320). При опускании крыловая
пластинка расположена горизонтально и движется вниз и вперед: возникает подъемная сила, удерживающая насекомое в воздухе. При движении вверх и назад крыло располагается вертикально, что создает про-пеллирующий эффект.
Количество ударов крыла в 1 с сильно варьирует у разных насекомых: от 5—10 (у крупных дневных бабочек) до 500—600 (многие комары); у
|
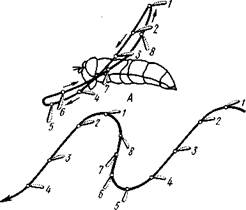
| Рис. 320. Схема траектории крыла насекомого. А — при стоячем полете мухи-журчалки; Б — при полете вперед (по Залесскому): 1—8 — восемь последовательных стадий положений крыла (сплошная черта — нижняя сторона крыла, пунктир — верхняя сторона) |
очень мелких комаров-мокрецов эта цифра достигает 1000 колебаний в 1 с. У различных представителей насекомых передние и задние крылья могут быть развиты в разной степени. Только у более примитивных насекомых (стрекозы) обе пары крыльев развиты более или менее одинаково, хотя и разнятся по форме. У жуков (отр. Жесткокрылые — Coleo-ptera) передние крылья изменяются в толстые и твердые надкрылья — элитры, которые почти не участвуют в полете и в основном служат для защиты спинной стороны тела. Настоящими же крыльями являются лишь задние крылья, которые в покоящемся состоянии спрятаны под надкрыльями. У представителей отряда клопов твердеет лишь основная половина
передней пары крыльев, вследствие чего эту группу насекомых нередко называют отрядом Полужесткокрылых. У некоторых насекомых, а именно у целого отряда двукрылых, развита лишь передняя пара крыльев, тогда как от задней остаются лишь рудименты в виде так называемых жужжалец.
|
Среди насекомых имеются и бескрылые, которые делятся на две различные группы. У одних отсутствие крыльев есть черта, присущая им искони и указывающая на примитивность их организации. Это все представители подкласса Entognatha и наиболее примитивные из открыточелюст-ных — отряд Thysanura (см. рис. 366). Среди остальных Ectognatha тоже встречаются формы, лишенные крыльев, — вши, блохи, некоторые мухи и т. п. Однако бескрылость в этом случае — свойство вторичное: это формы, утратившие крылья вследствие паразитического образа жизни или каких-либо иных причин.
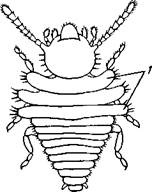
| Рис. 321. Нимфа Calotermes dilatatus с крыловидными расширениями (1) на всех сегментах груди (из Иммса) |
Вопрос о происхождении крыльев еще не вполне разрешен. В настоящее время одной из наиболее обоснованных представляется «паранотальная» гипотеза, согласно которой крылья возникли из простых неподвижных боковых выростов кожи — паранотумов. Такие выросты встречаются у многих члени-
стоногих (трилобиты, ракообразные), у многих ископаемых насекомых (см. рис. 362) и у некоторых современных форм (личинки термитов, некоторые богомолы, тараканы и т. д.; рис. 321). Переходом от ползающего к летающему послужил, быть может, лазающий образ жизни на деревьях, при котором насекомые, вероятно, часто делали прыжки с ветки на ветку, что способствовало дальнейшему развитию боковых выростов груди, которые вначале выполняли функции несущих плоскостей при парашютировании или планирующем полете. Дальнейшая дифференцировка и отчленение выростов от самого тела привели к выработке настоящих крыльев, обеспечивающих активный пропеллирующий полет.
Брюшко — последний отдел тела насекомых (см. рис. 307). Количество сегментов, входящих в его состав, варьирует у разных представителей класса. Здесь, как и в других группах членистоногих, выявляется четкая закономерность: чем ниже в эволюционном отношении стоят те или иные представители, тем более полным набором сегментов они обладают. И действительно, максимальное число брюшных сегментов мы находим у самых низших скрыточелюстных (отр. Protura), брюшко которых состоит из 11 сегментов и заканчивается отчетливо различимым тельсоном (см. рис. 363, А, Б). У всех остальных насекомых часть сегментов редуцируется (обычно один или несколько последних, а иногда и самый первый), так что общее число сегментов может сокращаться до 10, а у высших форм (некоторые перепончатокрылые и двукрылые) до 4-5.
|
Брюшко обычно лишено конечностей. Однако вследствие происхождения насекомых от форм, обладавших ножками на протяжении всего гомономно расчлененного тела, нередко сохраняются на брюшке рудименты конечностей или же конечности, изменившие свою первоначальную функцию. Так, у отряда Protura, низших представителей бескрылых насекомых, имеются маленькие конечности на трех передних члениках брюшка. Сохраняются рудименты брюшных конечностей и у открыто-челюстных. У тизанур на всех сегментах брюшка имеются особые придатки— грифельки, на которых, как на полозьях, при движении насекомого брюшко скользит по субстрату (см. рис. 366, А). Одна пара . грифель-
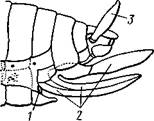
Рис. 322. Схема строения
яйцеклада насекомого
(из Бей-Биенко):
/ — половое отверстие. 2 —
створки яйцеклада, 3 — цер-
кн
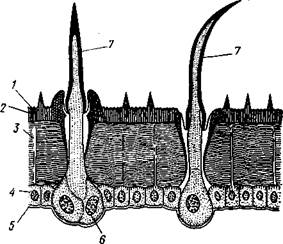
Рис. 323. Строение покровов насекомого (из Бей-Биенко) :
/— наружный слой кутикулы, 2 — средний слой кутикулы, 3 —внутренний слой кутикулы, 4 — гиподерма, 5 — базальная мембрана. 6 — клетка, образующая волосок, 7 — волосок
ков на заднем конце тела сохраняется и у тараканов. Очень широко распространены, особенно у более примитивных форм (тараканы, саранчовые и т. п.), церки — парные придатки последнего сегмента брюшка, также являющиеся видоизмененными конечностями. По-видимому, сходное происхождение имеют и яйцеклады (рис. 322), встречающиеся у многих насекомых и состоящие из трех пар вытянутых створок.
Покровы насекомых, как и всех остальных членистоногих, состоят из трех основных элементов — кутикулы, гиподермы и базальной мембраны (рис. 323). Кутикула выделяется клетками гиподермы, которая у скры-точелюстных насекомых часто превращается в синцитий. Кутикула насекомых трехслойна. В отличие от таковой ракообразных (с. 296) она несет наружный слой, содержащий липопротеинозые комплексы и препятствующий испарению воды из организма. Насекомые — сухопутные животные. Интересно отметить, что у водных и почвенных форм, обитающих в атмосфере, насыщенной парами воды, наружный слой или не выражен совсем, или развит очень слабо.
Механическую прочность кутикуле придают белки, задубленные фенолами. Они инкрустируют средний, основной слой.
На поверхности кутикулы находятся разнообразные, подвижно сочлененные с поверхностью тела выросты — тонкие волоски, чешуйки, щетинки. Каждое такое образование обыкновенно есть продукт выделения одной крупной гиподермальной клетки. Многообразие форм и функций волосков необычайно, они могут быть чувствительными, покровными, ядовитыми.
Окраска насекомых в большинстве случаев зависит от присутствия в гиподерме или в кутикуле особых красящих веществ — пигментов. Металлический блеск многих насекомых относится к числу так называемых структурных окрасок и имеет другую природу. Особенности строения кутикулы обусловливают появление ряда оптических эффектов, которые основаны па сложном преломлении и отражении световых лучей. Покровы насекомых имеют разнообразного значения железы; они бывают одноклеточными и многоклеточными. Таковы вонючие железы (на груди клопов), защитные железы (у многих гусениц) и т. д. Наиболее часто встречаются линочные железы. Их секрет, выделяемый во время линьки, растворяет внутренний слой старой кутикулы, не затрагивая вновь образующихся кутикулярных слоев. Особыми восковыми железами у пчел, червецов и некоторых других насекомых выделяется воск.
Мышечная система насекомых отличается большой сложностью и высокой степенью дифференциации и специализации отдельных ее элементов. Количество отдельных мышечных пучков часто достигает 1,5 — 2 тыс. Скелетные мышцы, обеспечивающие подвижность организма и отдельных его частей по отношению друг к другу, как правило, прикрепляются к внутренним поверхностям кутикулярных склеритов (тсргитам, стернитам, стенкам конечностей). По гистологическому строению почти все мышцы насекомых поперечнополосатые.
Мышцы насекомых (в первую очередь это относится к крыловым мышцам высших групп 'насекомых: перепончатокрылым, двукрылым и т. п.) способны к необычайной частоте сокращений — до 1000 раз в секунду. Это связано с явлением умножения ответа на раздражение, когда на один нервный импульс мышца отвечает несколькими сокращениями.
Богато разветвленная сеть трахейной дыхательной системы снабжает кислородом каждый мышечный пучок, что наряду с заметным повышением температуры тела насекомых при полете (за счет тепловой энергий, выделяемой работающими мышцами) обеспечивает высокую интенсивность обменных процессов, протекающих в мышечных клетках.
Пищеварительная система начинается небольшой ротовой полостью, стенки которой образуются верхней губой и совокупностью ротовых конечностей. У форм, питающихся жидкой пищей, она, по существу, заменена каналами (см. рис. 312,5), образующимися в хоботке и служащими для засасывания пищи и проведения слюны — секрета специальных слюнных желез. Стенки верхней части ротовой полости и следующей за ней трубчатой глотки соединяются со стенками головной капсулы с помощью мощных мышечных пучков. Совокупность этих пучков образует своеобразный мышечный насос, обеспечивающий продвижение пищи в пищеварительныю систему.
|
В заднюю часть ротовой полости, как правило, около основания нижней губы (максиллы II) открываются протоки одной или нескольких (до 3) пар слюнных желез. Содержащиеся в слюне ферменты обеспечивают начальные этапы процессов пищеварения. У кровососущих насекомых (муха цеце, некоторые виды комаров и т. п.) слюна часто содержит вещества, препятствующие свертыванию крови — антикоагулянты. В некоторых случаях слюнные железы резко меняют свою функцию. У гусениц бабочек, например, они превращаются в прядильные, которые вместо слюны выделяют шелковистую нить, служащую для изготовления кокона или для других целей.
Пищеварительный канал насекомых (рис. 324), начинающийся глоткой, состоит из трех отделов: передней, средней и задней кишок.
Передняя кишка может быть дифференцирована на несколько отличающихся по функциям и строению частей. Глотка переходит в пищевод, имеющий вид узкой и длинной трубки. Задний конец пищевода часто расширяется в зоб, особенно развитый у насекомых, питающихся жидкой пищей. У некоторых хищных жуков, прямокрылых, тараканов и т. д. за зобом помещается еще одно небольшое расширение передней кишки — жевательный желудок. Кутикула, выстилающая всю переднюю кишку, в жевательном желудке образует многочисленные твердые выросты в виде бугров, зубцов и т. п., способствующих дополнительному перетиранию пищи.
Далее следует средняя кишка, в которой происходит переваривание и всасывание пищи; она имеет вид цилиндрической трубки. При начале средней кишки в нее нередко впадает несколько слепых выпячиваний Рис- 324- Пищеварительная система
черного таракана (по Веберу): кишечника, или пилорических при- ;_м'юнные ж£иы. 2ЛищеволУ_аов.
Дата добавления: 2015-01-24; просмотров: 1656;