Ножки. 8 — яичник
Cirripedia (рис. 289). Такая циприсовидная личинка прикрепляется к
|
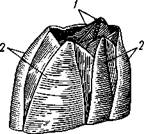
телу краба (чаще всего в месте сочленения суставов ножек), отбрасывает прочь грудь и брюшко и проникает внутрь хозяина в виде неправильной клеточной массы, которая питается за счет соков хозяина и медленно смещается вдоль кишечника краба в его брюшко. Разросшийся там паразит разрывает нежные брюшные покровы краба и выпячивается наружу в виде мешкообразной половозрелой саккулины.
| (по Дарвину): / — известковые пластинки крыши, 2 — пластинки доми- |
Отряд 5. Мешкогрудые (Ascothoracida).Небольшой отряд паразитических ракообразных с Рис 288 Морской же- организацией, более или менее измененной пара-лудь Balanus hammeri зитизмом. Это паразиты коралловых полипов и
иглокожих.
Наименее измененные паразитизмом формы — представители рода Sinagoga, живущие на колониях коралла Antipates и на теле морских лилий, могут еще ползать и даже плавать и сохраняют все характерные черты свободноживущего ракообразного. Самцы и самки их имеют сходное строение.
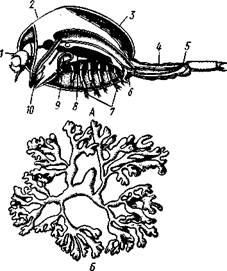
Сегментация и конечности хорошо развиты еще и у обитающего в половых сумках офиур Ascothorax (рис. 290, А), но их самки гораздо крупнее самцов. Напротив, у Denarogaster — эндопаразита морских звезд — произошли сильные изменения организации. Тело очень крупной (до 10 см) самки (рис. 290, Б) состоит из непарной срединной части, от которой в стороны отходят два сильно разветвленных, наподобие оленьих рогов, мантийных выроста. Непарная часть, представляющая собой собственно несегментированное туловище рачка, несет антеннулы и видоизмененные ротовые конечности. Грудные ножки отсутствуют.
|
| /J |
| 13 |
| 10 |
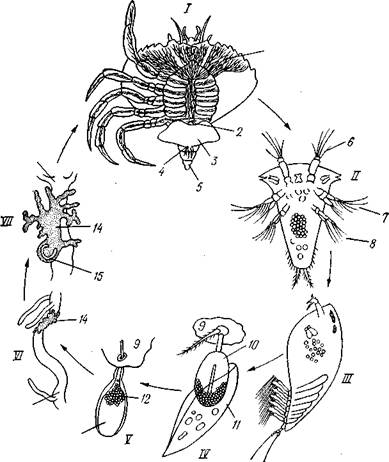
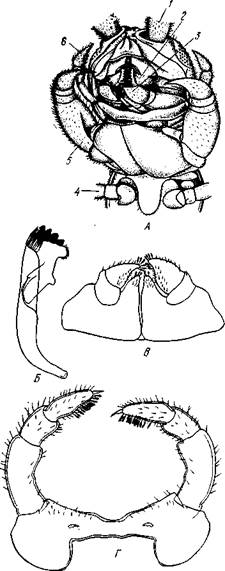
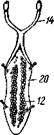
Рис. 289. Стадии развития Sacculina (из Арон и Грассе с изменениями). / — взрослая саккулина на брюшке краба; // — науплиус; III — ципри-совидная личинка; IV—она же, прикрепившаяся, у основания волоска на покровах краба (стадия кентрогон); V — внедрение в полость тела хозяина клеточной массы; VI — стадия внутренней саккулины на поверхности кишечника краба; VII — она же, образование мантии и мантийной полости:
1 — корневидные выросты саккулины, 2 — стебелек — основание корневой системы выростов, 3 — тело саккулины, 4 — отверстие мантийной полости, 5 — отогнутое брюшко краба. 6—антеннула, 7 — антенна, 8 — мандибула, 9 — кусок хитиновой кутикулы краба, 10 — кентрогон, II — отбрасываемый панцирь циприсовидной личинки, 12 — клеточная масса, мигрирующая внутрь краба, 13 — кишечник краба, 14 — стадия внутренней мигрирующей саккулины, 15 — зачаток мантийной
полости
В полости мантийных выростов располагаются яичники и ветви кишки. Там же помещаются и карликовые самцы Dendrogaster, вполне похожие на обычных ракообразных; тело самцов сегментированное, с конечностями и парой длинных мантийных выростов, в которые, как и у самок, проникают ветви кишечника и семенники.
Из яиц выходят науплиусы, которые покидают тело матери. У сильно измененных паразитизмом форм развитие продолжается в мантийных полостях самки, и в воду выходят личинки, по строению сходные с наиболее примитивными представителями мешкогрудых.
| 11—763 |
|
ПОДКЛАСС IV. РАКУШКОВЫЕ РАКИ (OSTRACODA)
Сюда относятся мелкие, часто различимые только под микроскопом морские и пресноводные ракообразные, отличающиеся крайней специализацией. Во-первых, тело их целиком заключено в двустворчатую раковину— разросшийся кара-паке. Во-вторых, туловище отличается крайним уменьшением числа сегментов. Остракоды — конечный этап очень древней эволюционной линии, совершенно независимой от подкл. Branchiopoda, которых они на первый взгляд напоминают.
| Рис. 290. Мешкогрудые раки. Л — Asco-thorax ophioctenis, самец; Б — Dendro-gaster dichotomus, самка (по Вагину): / — антеннула, 2—местооснования мантийной складки, 3 — мантия, 4 — брюшко, 5 — анус, 6 — кишка, 7 — грудные ножки, 8—нервная система (зачернена), 9 — максиллярная железа, 10 — ротовой конус |
Туловище остракод утратило всякие следы сегментации, но, если судить по числу ножек, грудь у них состоит только из 2—3 сегментов. Имеется науштиальный глаз, сложные глаза, как правило, отсутствуют. На голове, кроме того, расположены антеннулы, антенны, мандибулы и 2 пары максилл. Затем следуют 2 пары грудных ножек и, в некоторых случаях, еще пара так называемых чистилок, которые, вероятно, представляют видоизмененные ножки. Главным органом передвижения обычно служат антенны, но у некоторых пресноводных видов для плавания употребляются одновременно обе пары усиков. Грудные ножки играют существенную роль при ползании. Сердца и жаберных придатков у большинства остракод нет. Из яйца выходит науплиус, уже обладающий раковиной и представляющий, следовательно, хороший пример сдвига на раннюю стадию развития закладки органа, сильно развитого во взрослом состоянии.
Известно около 2000 видов ракушковых. В большинстве — это обитатели морей, но имеется и множество пресноводных видов, а южноафриканский Mesocypris terresfris живет даже в лесной подстилке влажного тропического леса.
Ракушковые питаются очень разнообразной пищей — растительной и животной, среди них есть даже прожорливые хищники. Они ведут плавающий (планктонный), ползающий и роющий образ жизни.
К самым обычным пресноводным формам у нас в СССР принадлежат Cypris pubera и Heterocypris reptans (см. рис. 256), встречающиеся массами в мелких водоемах и лужах. Размеры остракод колеблются у морских видов от 0,2 до 23 мм, а у пресноводных — от 0,3 до 7,3 мм.
Ископаемые остракоды встречаются начиная с кембрия. Ввиду разнообразия и многочисленности они имеют известное значение при определении местонахождений нефти наряду с фораминиферами.
ПОДКЛАСС V. ВЫСШИЕ РАКИ (MALACOSTRACA)
В противоположность остальным ракообразным подавляющее большинство представителей подкласса Malacostraca обладает постоянным
числом сегментов: 4 головных, 8 грудных и 6 брюшных (исключение составляют только тонкопанцирные раки отряда Leptostraca; см. ниже). Голова или образует цельную головную капсулу — сложную голову, в состав которой кроме акрона и 4 головных сегментов входит первый сегмент груди (отр. Amphipoda, Isopoda), или представлена протоцефало-ном (акрон +сегмент антенн). В последнем случае челюстные сегменты головы сливаются с несколькими или со всеми сегментами груди в особый отдел, называемый челюстегрудью. Брюшко снабжено 6 парами
|
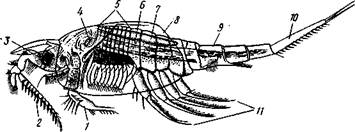
Рис. 291. Небалия Nebalia geoffroyi, самец (по Клаусу):
/ — антенна, 2 — антеннула, 3 — глаз, 4 — поперечная мышца, 5 — грудь,
6 — семенник, 7 — сердце, 8 — двускатный панцирь, 9—брюшко, 10 —
тельсон, // — брюшные ножки
конечностей. Органами выделения во взрослом состоянии, как правило, служат антеннальные железы. Половые отверстия лежат у самки на 6-м, у самца на 8-м грудном сегменте. В развитии характерна личинка зоеа.
Русское название подкласса не совсем удачно, так как некоторые черты строения, например развитие на всех брюшных сегментах двувет-вистых конечностей, явно примитивнее, чем у других ракообразных. Вероятно, высшие ракообразные в процессе эволюции развились как самостоятельная ветвь независимо от других подклассов; очевидно, в каждом из подклассов Crustacea сохранились свои специфические примитивные особенности строения и развития.
К подклассу Malacostraca, объединяющему свыше 14000 видов, относится 14 отрядов, из которых ниже рассматриваются лишь главные.
Отряд 1. Тонкопанцирные (Leptostraca).Небольшой, состоящий всего из 8 видов отряд мелких морских раков, обладающих некоторыми чертами низкой организации. Leptostraca имеют 7 (не 6) сегментов брюшка; голова, грудь и часть брюшка покрыты двускатным панцирем, между половинами которого имеется поперечная мышца (как у Ostracoda). Во взрослом состоянии обладают не только антеннальными, но и слегка редуцированными максиллярными железами. Представитель Nebalia (рис. 291).
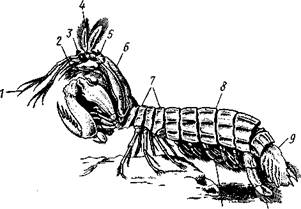
Отряд 2. Ротоногие (Stomatopoda).Небольшой, но своеобразный отряд морских раков. Тело вытянутое (до 34 см длиной), с очень длинным и мощным брюшком. Имеется протоцефалон. Четыре первых грудных сегмента входят в состав челюстегруди. Передние 5 пар (в особенности же 2-я пара) грудных ног изменены в хватательные конечности. Последний членик 2-й пары ног сплющен с боков в виде зазубренного лезвия и может, как перочинный нож, вкладываться в особый желоб предпоследнего членика. Жабры развиты на грудных и особенно брюшных конечностях.
Взрослые ротоногие раки большей частью ведут роющий образ жизни на дне моря, личинки встречаются в планктоне. Stomatopoda живут
| 11* |
преимущественно в теплых морях. Число известных до сих пор видов около 170. Представитель — рак-богомол (Squilla oratoria; рис. 292) до 20 см длины. В Средиземном море, а также в Тихом и Индийском океанах существует промысел некоторых крупных ротоногих, употребляемых в пищу.
// V/7
Рис. 292. Рак-богомол Squilla oratoria (из Бирштейна): / — антеннулы, 2 — антенны, 3—• глаза, 4 — наружные чеш;н антенн, 5 — первичная голова, 6—карапакс, 7 — грудные сегменты, 8 — брюшко, 9 — тельсон, 10 — последняя пара брюшных ног, 11 —- брюшные ноги
Отряд 3. Мизиды(Mysidacea). Ракообразные, внешне напоминающие небольших креветок (с. 328), но сходство это поверхностное и вызвано сходным образом жизни, именно приспособлением к плаванию. Длина тела в среднем от 10 до 20 мм. Известно около 500 преимущественно морских, реже пресноводных видов.
Имеется протоцефалон. В состав челюстегруди входит не более трех передних грудных сегментов. Одна передняя пара грудных ножек превращена в ногочелюсти. Все грудные конечности двуветвисты. Жабр нет, газообмен совершается через стенку карапакса. Питаются мизиды мелкими частицами детрита, которые отфильтровывают щетинками обеих нижних челюстей и ногочелюстей.
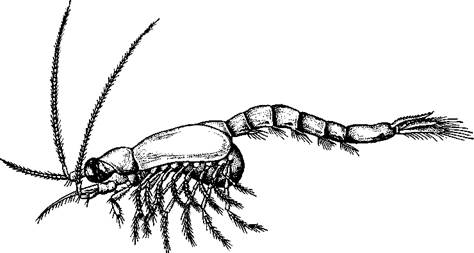
Рис. 293. Расщепленноногий рачок Mysis relicta (по Сарсу)
Самка вынашивает яйца в выводковой сумке, расположенной на груди. Из сумки выходит молодь, мало отличающаяся от взрослых животных.
К обычным представителям мизид относится Mysis rslicta (рис. 293), обитающий в холодных и чистых озерах северных областей европейской части СССР, в Северной Европе и Северной Америке. Практическое значение мизид велико — они составляют существенную долю в пищевом рационе некоторых промысловых и искусственно разводимых рыб.
Отряд 4. Кумовые (Сшпасеа).Небольшие (от 10 до 35 мм) ракообразные, в общем близкие к мизидам, но ведут роющий образ жизни. Переднебоковые углы карапакса
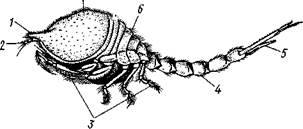
Рис. 294. Самка кумового рачка Diastylis goodsiri (по Сарсу):
/ — передний угол карапакса, 2 — антеннула, 3—грудные ниги, 4 — брюшко, 5 — тельсон, 6 — грудь, 7 — карапакс
вытянуты вперед, сближены и несут небольшие боковые отверстия, сообщающие пространство под панцирем с наружной средой. В остальных местах края карапакса плотно прилегают к телу. Зарываясь в грунт, рачок выставляет наружу только передний конец тела с отверстиями карапакса, через которые вода проникает под панцирь и омывает дыхательные полости.
В состав челюстегруди входят 3 передних сегмента груди, конечности которых превращены в ногочелгости. Грудные ножки большей частью двуветвисты, брюшные конечности частично не развиты. Сложные глаза рудиментарны или отсутствуют. Яйца вынашиваются самкой в выводковой сумке на груди. Развитие без метаморфоза.
Кумовые — преимущественно морские обитатели, лишь немногие живут в пресных водах. Чисто пресноводная форма Lamprops corroensis живет в озерах и реках Дальнего Востока. К. обычным морским представителям относятся роды Cumopsis и Diaftylis (рис. 294). Кумовые — излюбленная пища некоторых рыб.
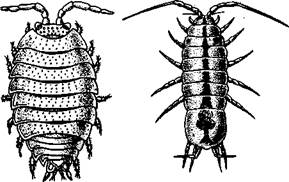
| Рис. 295. Равноногие раки Isopoda. A — мокрица РогсеШо; Б — водяной ослик Asellus aquaiicus (чз Догеля) |
Отряд 5. Равноногие (Isopoda).Большая (4500 видов) процветающая группа ракообразных, отличающаяся высокой пластичностью организа-
шш; среди равноногих немало морских, пресноводных, а также наземных и паразитических форм.

|
| Рис. 296. Самка паразитического равноногого рака Cancricepon elegans (из Бирштейна): / — выводковая сумка, 2 — грудные ножки, 3 — грудь, 4 — брюшные ножки, 5 — уроподы, б — брюшко, 7 — карликовый самец, сидящий на самке |
Тело обычно уплощено дорзовентрально (рис. 295). Размеры тела колеблются от 1 мм до 5 см, только глубоководный морской Bathynomus достигает 27 см. У равноногих раков сложная компактная голова, в состав которой кроме акрона и головных сегментов входят еще 1—2 сегмента груди. Конечности последних превращены в но-гочелюсти. На голове расположены большие фасеточные глаза. Карапакс отсутствует; грудные сегменты несут одноветвистые ходильные ножки. Брюшко короче груди, в большинстве случаев все брюшные сегменты или часть их срастаются с анальной лопастью. Пять пар передних брюшных ног служат для дыхания; они состоят из короткого основания и двух широких листовидных жаберных ветвей, которые направлены назад
и прилегают друг к другу, как листы книги. Экзоподиты одной пары брюшных ног образуют прочную крышку, покрывающую все жаберные листочки. Такое строение дыхательного аппарата позволило некоторым представителям Isopoda приспособиться к жизни на суше. Примером могут служить мокрицы, дышащие кислородом, растворенным в тонком слое влаги, покрывающей жаберные листочки. Часть мокриц, впрочем, дышат атмосферным воздухом, у таких форм на экзоподитах передних брюшных ног имеется глубокое впячивание покровов, от которых отходят слепо замкнутые на концах дыхательные трубочки, называемые псевдотрахеями.
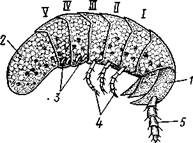
Среди паразитических изопод имеются временные (Aega) или постоянные (Cymothoa, Livoneca) эктопаразиты рыб. Некоторые формы паразитируют на ракообразных. Таков, например, Cancricepon (рис. 296), обитающий в жаберной полости креветок и крабов.
Яйца у нзопод развиваются в выводковой камере на груди самки до личиночной стадии, называемой манкой. Манка отличается от взрослых животных главным образом недоразвитыми последними и грудными ногами, но у паразитических форм имеет ряд специальных приспособлений к подвижной активной жизни, связанной с поисками нового хозяина. К широко распространенным представителям отряда принадлежит пресноводный водяной ослик — Asellus aquaticus (см. рис. 295, Б), обычны мокрицы — РогсеШо (см. рис. 295, Л) и Oniscus.
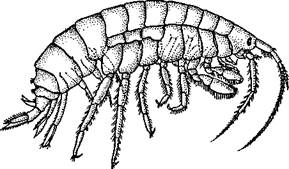
Отряд 6. Разноногие, или бокоплавы (Amphipoda)..По числу видов (4500) этот отряд не уступает предыдущему. Большинство бокоплавов обитает в море, среди них имеются как донные, в том числе роющиеся в грунте или живущие в защитных трубках, так и планктонные виды. Многие населяют пресные воды, немногие ведут паразитический образ жизни.
Тело бокоплавов большей частью сжато с боков (рис. 297). Голова, как и у равноногих, цельная, к ней присоединены 1—2 грудных сегмента. Глаза фасеточные. Карапакс отсутствует. Ножки всех грудных сегментов
|
устроены различно, некоторые вооружены хватательными крючками и почти все несут листовидные жаберные пластинки. Брюшные конечности хорошо развиты, три передние пары двуветвисты, снабжены плавательными щетинками и служат для плавания, задние пары направлены назад и вместе с тельсоном служат для прыгания. В период размножения у самок образуется на груди выводковая камера, в которой вынашиваются развивающиеся яйца. Из камеры выходит молодь, строением не отличающаяся от взрослых. Большинство бокоплавов
| Рис. 297. Бокоплав Gammarus lacustris (по Сарсу) |
всеядно, немало хищников, которые преобладают среди планктонных форм. Паразитами являются китовые вши — Cyamidae, грызущие кожу хозяина.
Среди представителей Amphipoda заслуживают
упоминания морские блохи — Gammarus и Anisogammarus, массами населяющие приливно-отливную зону многих морей. Особенно богато бокоплавами дно материкового склона северных морей. Так, в Чукотском море на 1 м2 дна обитает около 40 000 особей. К обычным пресноводным бокоплавам принадлежит озерный бокоплав — Gammarus lacustris, широко распространенный в северном полушарии (рис. 297). В Байкале имеется фауна нигде более не встречающихся бокоплавов (240 видов).
Практическое значение амфипод довольно велико, поскольку они составляют излюбленный корм различных рыб. В связи с этим некоторые пресноводные бокоплавы были перенесены и акклиматизированы в ряде озер и водохранилищ.
Отряд 7. Эуфаузиевые (Euphausiacea)—небольшой отряд высших ракообразных, насчитывающий лишь около 80 видов. Это планктонные обитатели моря, внешне похожие на небольших креветок (с. 328). Характеризуются наличием протоцефалона и че-люстегруди, в состав которой вошли все грудные сегменты, и развитием карапакса. Эуфаузиевые, как и креветки, имеют ряд сходных приспособлений к плаванию в толще воды. Однако Эуфаузиевые легко отличимы от креветок наличием свободных, не прикрытых карапаксом жабр, сидящих на основаниях грудных ножек (рис. 298). Последние двуветвисты и в отличие от десятиногих раков не образуют ногочелюстей и служат только для плавания.
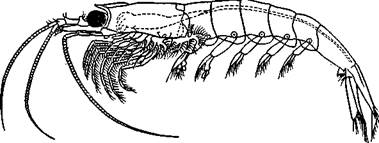
Рис. 298. Эуфаузиевый рак Euphausia pellucida (по Сарсу)
Характерны для эуфаузиевых хорошо развитые фасеточные стебельчатые глаза и органы свечения — фотофоры, особенно у глубоководных форм. Обычно фотофоров бывает 10 пар, расположенных на глазных стебельках, грудных и брюшных сегментах. Эуфаузиевые — хорошие пловцы, передвигающиеся главным образом при помощи сильных брюшных ног. Размеры тела колеблются от 7 до 96 мм.
Самка откладывает яйца в воду или прикрепляет их к брюшным ножкам. Из яйца выходит науплиус.
Эуфаузиевые массами размножаются в некоторых районах моря, где служат пищей различным морским млекопитающим и рыбам. Так, в антарктических водах скопления усатых китов связаны с зонами массового размножения так называемого криля — Euphausia superba. В Баренцевом море массовый вид Thysanoesia raschii — пища сельди, морского окуня, трески и других промысловых рыб.
Отряд 8. Десятиногие (Decapoda).Этот отряд объединяет крупных и во многих отношениях наиболее высокоорганизованных ракообразных. Имеется первичная голова — протоцефалон, несущая две пары усиков и стебельчатые глаза. Все сегменты груди сливаются с челюстными сегментами головы и покрыты карапаксом. Передние три пары грудных ног превращены в ногочелюсти! Первая пара ходных ног большей частью в виде клешней. Грудные конечности у более примитивных форм двувет-вистые, гребные, у большинства же одноветвистые, ибо экзоподит исчезает. Жабры располагаются на грудных конечностях, частью же на боках самого тела. Очень разнообразны форма и строение брюшка у десятиногих. В одних случаях брюшко большое, длинное, с хорошо развитыми ножками, которые служат для плавания. Последние, однако, у ряда форм сильно уменьшаются в размерах и активного участия в движении уже не принимают. В других случаях брюшко теряет часть конечностей, становится мягким и асимметричным (раки-отшельники). Наконец, крабы обладают очень небольшим, симметричным, подогнутым впереди брюшком, которое несет рудиментарные конечности.
К десятиногим принадлежит свыше 8500 видов. Они распространены очень широко, встречаясь на всех глубинах морей и океанов. Особенно разнообразна фауна десятиногих на мелководье тропических морей. К пресноводным формам относятся речные раки и некоторые крабы и креветки. Некоторые виды крабов и раков-отшельников перешли к наземному образу жизни.
Самки десятиногих прикрепляют яйца к брюшным ножкам и вынашивают их до вылуплсния молоди. Из яйца большей частью выходит личинка, резко отличающаяся от взрослого животного, например у крабов и раков-отшельников — зоеа, а у омаров — мизидная стадия. Только у некоторых низших креветок первая личиночная стадия представлена науплиусом. Для пресноводных и глубоководных морских форм характерно прямое развитие, когда из яйца выходит миниатюрное, почти сформированное животное.
Практическое значение десятиногих немаловажно, ибо многие из них представляют ценный продукт питания. Важное промысловое значение имеют речные раки (Potamobius), которые в СССР употребляются как пищевой продукт и являются ценным предметом экспорта, омары (Но-marus), лангусты (Palinurus), креветки (Crangon, Pandalus), а также многие крабы (Cancer, Callinectes). В СССР высоко развита крабокон-сервная промышленность, использующая «камчатского краба» — Рага-lithodes catntschatica.
Подотряд 1. Natantia объединяет наиболее примитивные формы десятиногих, обладающих хорошо развитым длинным брюшком, несущим конечности, используемые для плавания. Типичные представители этой группы — разнообразные креветки: Pandalus (рис. 229,Л), Crangon и др.
| со 1чЭ «С |
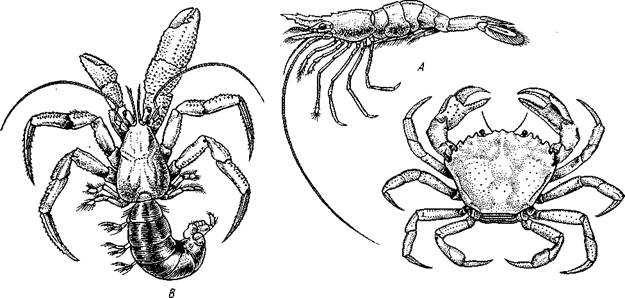
Рис. 299. Представители десятиногих ракообразных. А — креветка Pandalus borealis; Б — краб Carcinus тае-nas; В — рак-отшельник Pagurus bernltardiis (из Бирштейна)
Подотряд 2. Reptantia. По числу представителей значительно больший, включает формы, отличающиеся как по строению, так и по биологии. Общим признаком является то, что они не используют для плавания брюшные конечности. Последние развиты значительно слабее, чем у представителей подотр. Natantia, часто рудиментарны, а их количество уменьшено.
Reptantia подразделяются на несколько «отделов». Типичные представители отдела Palinura — лангусты (Palinurus)—довольно крупные морские ракообразные, без клешней. Широко известны и формы, относящиеся к отделу Astacura, — омары (Homarus), пресноводные речные раки (Potamobius; см. рис. 252) и др. У них, так же как и у лангустов, длинное симметричное брюшко с небольшими двуветвистыми конечностями, но первая пара ходных ног несет мощные клешни. Отдел Anomura, или неполнохвостые, включает разнообразных раков-отшельников (например, Pagurus; рис. 299,5), прячущих свое мягкое асимметричное брюшко в пустые раковины брюхоногих моллюсков. Некоторые неполнохвостые перешли к жизни на суше: наземный рак-отшельник—СоепоЫ-1а, краб-разбойник — Birgus latro. Последний представляет большой интерес как форма, более или менее хорошо приспособленная к наземному образу жизни. Краб-разбойник обитает на некоторых островах Тихого и Индийского океанов вдали от моря, роет неглубокую норку, которую покидает только ночью. Питается маслянистыми плодами тропических растений. В период вылупления личинок из яйца самка уходит в море. Вышедшие личинки в течение нескольких месяцев живут в планктоне, а затем опускаются на дно. Здесь они забираются в пустые раковинки моллюсков и становятся очень похожими на типичных раков-отшельников. В таком виде они и выходят на сушу, где через некоторое время оставляют раковину и превращаются во взрослых крабов-разбойников.
К этому же отделу относится камчатский краб — Paralithodes camlschatica (рис. 300), внешне похожий на настоящих крабов (с. 330). Он никогда не использует раковины моллюсков, однако его маленькое подогнутое вперед брюшко сохраняет асимметричное строение.

|
| Рис. 300. Камчатский краб Paralithodes camtschatica. В размахе ног самец достигает 1,5 м при массе до 7 кг (по Догелю) |
Представители отдела Brachyura (короткохвостые, или крабы) отличаются маленьким, подогнутым под грудь симметричным брюшком, короткими усиками и широким панцирем. Сюда относятся морские промысловые крабы Carcinus (см. рис. 299,Б), Cancer и многие другие, а также некоторые пресноводные и даже наземные формы.
ПОДТИП ТРАХЕЙНЫЕ (TRACHEATA)
Подтип трахейных включает наземных членистоногих, дышащих при помощи трахей. Подавляющее большинство трахейных обладает хорошо обособленной слитной головой, состоящей из акрона и четырех сегментов. Конечности первого сегмента, соответствующие антеннам II раков, у всех Tracheata отсутствуют, а сам сегмент, называемый вставочным или интеркалярным, может подвергаться частичной редукции. Придатки головы трахейных представлены одной парой усиков, или ан-теннулами, и тремя парами ротовых конечностей — верхними челюстями (мандибулами) и первыми и вторыми нижними челюстями (максилла-ми). Сегментарный состав туловища и его подразделение па отделы сильно варьируют в пределах группы. Будучи сухо-воздушными организмами, Tracheata приобрели ряд морфофизиологических адаптации к существованию в условиях дефицита влаги.
Подтип трахейных включает два класса: Многоножки (Myriapoda) и Насекомые (Insecta).
КЛАСС I. МНОГОНОЖКИ (MYRIAPODA)
Класс Myriapoda включает около 10000 видов исключительно наземных, иногда довольно крупных членистоногих. Червеобразное, вытянутое тело многоножек подразделяется на два отдела: слитную голову и чле-
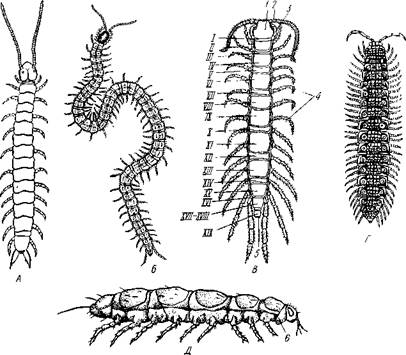
Рис. 301. Различные многоножки (из Беклемишева): А — Scolopendrella immaculata (подкл. Symphyla); Б — Pachimerium ferrugineum (подкл. Chilopoda); В — Lithobius forficatus (подкл. Chilopoda); Г — Polydesmus complanatus (подкл. Diplopoda); Д — Pauropus silvaticus (подкл.
Pauropoda):
1 — голова, 2 — антенна, 3 — ногочслюсть, 4 — туловище, 5 — анальное отверстие, I—XIX туловищные сегменты, 6 — шейный сегмент, гомологичный сегменту мак-
снлл II
нистое туловище, которое часто состоит из значительного числа снабженных конечностями сегментов.
Все многоножки объединяются в 4 подкласса: симфилы (Symphyla), пауроподы (Pauropoda), двупарноногие, или кивсяки (Diplopoda), и гу-боногие (Chilopoda).
Строение и физиология.Тело многоножек (рис. 301) состоит из резко
|
отделенных друг от друга сегментов, число которых варьирует в широких пределах: от 18 у симфил (подкл. Symphyla) и 14 у пауропод (подкл. Pauropoda) до 181 у некоторых губоногих (подкл. Chilopoda).
Голова Myriapoda ясно обособлена от туловища. Она включает акрон и слившиеся с ним 4 (подкл. Symphyla и Chilopoda) или 3 (подкл. Pauropoda и Diplopoda) первых сегмента тела. Во втором случае последний головной сегмент остается свободным и называ-
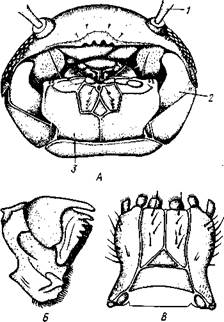
Рис. 302. Ротовой аппарат Lithobius for-ficatus (Chilopoda) (из разных авторов). А — внешний вид головы; Б — мандибула— верхняя челюсть; В — максиллы I; Г — максиллы II:
1 — основание усика. 2 — мандибула, 3 — максилла I,
4 — основание туловищной ножки, 5 — ногочелюсть,
6' — максилла II
Рис. 303. Ротовой аппарат Diplopoda (из разных авторов). А — внешний вид головы; 5 — мандибула; В — гнатохилярий:
/ — основание усика, 2 — мандибула, 3 — гнатохилярий
ется «-шейным». Эта особенность строения головы части многоножек справедливо рассматривается как примитивный признак.
Голова несет усики и ротовые конечности: верхние челюсти — мандибулы, или жвалы, и нижние челюсти — максиллы, которых соответственно числу сегментов, вошедших в состав головы, может быть одна или две пары.
Усики, или антенны, многоножек отвечают антеннулам (антеннам I) раков и относятся к акрону. Они более или менее длинные, тонкие, разбиты на членики и большей частью неветвистые. По-видимому, они служат органами не только осязания, но и обоняния. Конечности, соответ ствующие антеннам II раков и принадлежащие первому сегменту тела4 у многоножек редуцированы. Но их сегмент, называемый пнтеркаляр-ным, имеется. Прочие головные конечности превращены в ротовые органы и гомологичны соответствующим ротовым частям раков. Строение их неодинаково в разных подклассах многоножек. У представителей подклассов Symphyla и Chilopoda (рис. 302) рот спереди прикрыт хити-низированной складкой покровов — верхней губой; последняя по происхождению не имеет ничего общего с конечностями. Мандибулы, конечности второго сегмента, состоят из двух коротких жевательных пластинок с зазубренным внутренним краем. Первая и вторая пары нижних челюстей (конечности III и IV сегментов соответственно) у большинства представителей состоят из основания, на котором сидят членистый челюстной щупик и нерасчлепенные жевательные лопасти. И щупик и лопасти могут частично редуцироваться.
У представителей подклассов Pauropoda и Diplopoda (рис. 303) за верхней губой и парой мощных зазубренных мандибул располагается всего одна непарная пластинка — гнатохилярий (gnathochilarium) довольно сложного строения. История развития показывает, что она закладывается в виде парного зачатка и соответствует первой паре нижних челюстей губоногих. Конечности шейного сегмента редуцировались1.
За головой следует большей частью однородно построенное туловище. Однако строго гомономная сегментация бывает выражена только у самых примитивных форм (см. рис. 301,Б). В процессе эволюции характер сегментации заметно меняется. У некоторых многоножек (ряд губоногих) часть туловищных сегментов заметно уменьшается в размерах (см. рис. 301,6). При этом уменьшенные и нормальные сегменты довольно правильно чередуются. Иная картина наблюдается у представителей подкласса Diplopoda, у которых происходит попарное слияние большей части сегментов (кроме первых четырех, включая и «шейный» сегмент). Каждый такой двойной сегмент соответственно несет не одну, а две пары конечностей (рис. 304).
Подобные отступления от исходной примитивной гомономности не ведут, однако, к подразделению туловища на тагмы. Лишь у кивсяков первые туловищные сегменты, несущие по одной паре конечностей (рис. 304) 'и тем отличающиеся от прочих, вместе с безногим «шейным» сегментом иногда обозначаются как «грудные», а следующие за ними двойные сегменты — «брюшные».
Однородность туловищных сегментов многоножек определяет и сходство строения их конечностей, которые имеют вид простых ходных ножек, состоящих из одного ряда члеников и заканчивающихся коготком. Примеры их функциональной и морфологической дифференциации не-
1 Существует и другая точка зрения, согласно которой гнатохилярий Diplopoda образуется за счет максиллИ. Шейный сегмент в этом случае рассматривается как первый туловищный сегмент.
многочисленны. Так, у губоногих характерно превращение ножек первого туловищного сегмента в ногочелюсти (см. рис. 302,Л), играющие главную роль в захвате и убивании добычи. Эта пара ножек сильно увеличивается в размерах и обладает крайне утолщенным основным члеником, тогда как концевой членик сильно заострен и загнут в виде крюч-
|
| /// |
| /7- |
Рис. 304. Передний и задний концы тела с левой стороны самца кивсяка Schizophyllum sabulosum (по Иванову):
/ — голова, 2—глазное поле, 3—антенна, 4—обонятельные органы, 5 —верхняя губа, 6 — верхняя челюеть, 7 — щека, 8 — видоизмененная ножка 1-го туловищного сегмента, сместившаяся вперед. 9 — ножка 2-го туловищного сегмента, сместившаяся вперед, 10 — место полового отверстия. И — ножка 3-го сегмента, 12 — ножки двойных сегментов, 13 — тельсон, 14 — «молодые» сегменты зоны роста, /Л — сегменты, слившиеся в двойной сегмент, 16 — отверстия ядовитых желез, 17—«шейный» сегмент, соответствующий сегменту максплл И; I — 1П — сегменты «груди»; IV. V, VI н т. д. — сегменты «брюшка»
ка. В основании конечности залегает ядовитая железа, проток которой открывается на конце крючка. Выделяемый яд сильно действует на членистоногих и позвоночных. Укус пальца крупной сколопендрой (Scolo-pendra) приводит к временному опуханию всей руки. Некоторые пары ножек, принимающие участие в копуляции, несколько изменяются в строении и называются гоноподиями.
Тело покрыто хитиновой, иногда пропитанной известью кутикулой, выделяемой однослойным гиподермальным эпителием. Последний довольно богат одноклеточными и многоклеточными кожными железами, среди которых особенно интересны защитные железы кивсяков. Они помещаются на спинной стороне части туловищных сегментов и открываются наружу оборонительными отверстиями, из которых выпрыскивается секрет. У разных видов подкласса Diplopoda секрет необычайно варьирует по виду и химическим свойствам. Так, секрет Spirobolus едок и окрашивает кожу человека в темный цвет. Polyzonium rosalbum выделяет млечную жидкость, имеющую запах и жгучий вкус камфоры. Тропическая Fontarla содержит в железах свободную синильную кислоту и пахнет горьким миндалем.
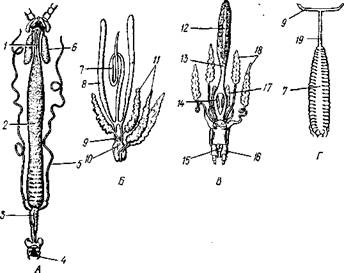
Пищеварительная система многоножек (рис. 305) имеет вид прямой трубки; лишь в области задней кишки пищеварительный канал образует петлевидный изгиб.
Рот лежит на брюшной стороне головы между ротовыми конечностями и ведет в переднюю кишку, часто называемую пищеводом. С начальным отделом пищеварительной системы связаны слюнные железы. Кив-сяки обладают тремя парами желез, открывающимися самостоятельными протоками в ротовую полость и у основания гнатохиляриума. Ввиду образования их из мезодермы эти железы считают видоизмененными целомодуктами. У губоногих 3—5 пар слюнных желез с самостоятель-
ными протоками, открывающимися в ротовую полость или по бокам рта. Они, по-видимому, чисто эктодермального происхождения, т. е. представляют модифицированные кожные железы. Слюнные железы, открывающиеся на второй паре челюстей, приравнивают к прядильным желе-
|
|
Рис. 305. Внутренние органы многоножек. А—пищеварительная система Lithobius fofficalus (Chilopoda) (по Плате); Б — мужская половая система Chilopoda (из Шауфлера); В — женская половая система Chilopoda (из Шауфлера); Г—мужская половая система Diplopoda (по Фабру); Д —
женская половая система Diplopoda (по Фабру):
Пищевод, 1 — средняя кишка, 3 — задняя кишка, 4—анус, 5 — мальпигиевы сосуды, в — слюнная железа, 7 — семенник, 8 — семенной пузырек, 9 — парный участок семяпровода, 10 — семяизвергательный канал, // — придаточные железы, 12 — яичник, 13 — непарный участок яйцевода, 14 — парный участок яйцевода, 15 — половое отверстие, 16 — половая ножка, 17 — семяприемник, 18 — придаточные железы. 19 — семяпровод, 20 — вскрытый половой мешок
зам личинок насекомых, отверстия которых помещаются на той же паре ротовых конечностей.
Средняя кишка служит местом переваривания и всасывания пищи. Задняя кишка короткая.
Представители подкласса Diplopoda растительноядны и питаются главным образом гниющими листьями, растительными остатками, древесной трухой и т. п. Губоногие — хищники, питающиеся насекомыми.
Выделительная система. На границе между средней и задней кишками в кишечник впадают 1 или 2 пары (последнее у Chilopoda) длинных слепозамкнутых на свободном конце трубок — мальпигиевых сосудов (рис. 305,Л). В эпителии сосудов и их просвете скапливаются конкреции мочевой кислоты; последняя у многоножек, так же как и у насекомых (с. 352), представляет собой главный продукт выделения. Кроме мальпигиевых сосудов выделительную роль играют и другие образования, прежде всего лимфатические железы в виде неправильных клеточных тяжей, расположенных либо вдоль мальпигиевых сосудов, либо вдоль брюшного кровеносного сосуда, или вдоль брюшной нервной цепочки. Они служат для захвата и накопления твердых продуктов выделения и фагоцитнруют впрыснутые в полость тела твердые частицы (например, порошка туши или кармина). Кроме того, участие в выделении прини-
мает жировое тело. Полость тела — миксоцель — многоножек во многих местах заполнена неправильными тяжами и скоплениями клеток, причем эти скопления ограничены тонкой собственной оболочкой. Совокупность таких скоплений и называется жировым телом. В клетках его наблюдаются многочисленные капли жира, а также конкреции мочевой кислоты. Жировое тело служит не только для накопления запасного питательного материала, но и для выделения (мочевая кислота).
Нервная система состоит из головного мозга, окологлоточных кон-нективов и брюшной нервной цепочки. Головной мозг имеет довольно сложное гистологическое строение, свидетельствующее, что голова многоножек образована из сегментов, превышающих по числу количество пар головных придатков. В мозге помимо пары ганглиев, посылающих нервы к антеннам, имеются еще парные скопления нервных клеток, отвечающие вставочному (интеркалярному) сегменту.
Брюшная цепочка состоит из расположенного в голове подглоточного ганглия, который инпервирует все ротовые конечности, и из длинного ряда туловищных ганглиев, хорошо обособленных и сидящих на общем парном продольном нервном стволе. Па каждый сегмент приходится обычно один парный ганглий. У Diplopoda такое устройство наблюдается лишь в четырех передних сегментах, прочие же содержат по два расположенных один за другим ганглия, чем доказывается сложный состав этих сегментов.
Органами осязания и обоняния служат антенны, усаженные чувствительными волосками, чувствительными колбочками и т. п. Кроме того, по бокам головы, между основаниями антенн и глазами, лежат два темсшваровых органа чувств (по-видимому, хеморецепторы). Это или подковообразные ямки, на дне которых имеются валики из чувствительных клеток, или же скопления чувствительных клеток, помещающиеся под покровами головы в глубине длинных узких каналов. Темешваровы органы иннервируются от головного мозга. Громадное большинство многоножек снабжено глазами, их может быть 2, 4 или много. Глаза сидят по бокам головы и носят характер одиночных, просто устроенных глазков. Только у мухоловок (Scutigera) на голове два больших скопления глазков, так тесно сближенных, что они соприкасаются друг с другом и напоминают сложные фасетированные глаза насекомых. Зрительные способности многоножек невелики. Многоножки предпочитают затененные участки.
Органы дыхания представлены трахеями — тонкими воздухоносными трубочками эктодермального происхождения, возникающими как глубокие впячивания покровов. Стенки трахей выстланы продолжением наружной кутикулы, которое на всем протяжении трахейной трубки образует спиральное утолщение, не дающее трахеям спадаться. Трахеи начинаются парными дыхальцами, или стигмами, лежащими на брюшной стороне сегментов туловища. Как исходная форма трахейной системы должна рассматриваться такая, у которой каждый туловищный сегмент несет пару стигм, а каждая стигма ведет в обособленный пучок тонких трахейных трубочек. Ближе всего к этой схеме стоит подкласс Diplopoda, у представителей которого почти все туловищные сегменты снабжены независимыми друг от друга парными пучками неразветвленных трахей. В связи с двойным характером туловищных сегментов последние несут не по 1, а по 2 пары дыхалец. У большинства видов подкласса Chilopoda стигмы расположены на туловище через сегмент, а у некоторых форм (например, Scutigera) животное имеет всего 7 пар стигм, но сама трахейная сеть развита у губоногих гораздо сложнее. Трахеи у некоторых из них сильно ветвятся, и между трахейными пучками соседних сегмен-
тов и одного и того же сегмента (правым и левым) устанавливаются сообщения в виде продольных и поперечных перемычек. Концевые веточки трахеи многоножек оплетают все внутренние органы. Смена воздуха в трахеях происходит вследствие изменения объема тела при сокращении и расслаблении мускулатуры.
Кровеносная система довольно хорошо развита, кроме сердца есть система периферических кровеносных сосудов. Сердце в виде нежной прозрачной трубки тянется над кишкой вдоль всего туловища и сзади замыкается слепо или же продолжается в два коротких сосуда, теряющихся в мускулатуре. Сердце соответственно сегментам поделено на камеры: каждая камера имеет две остии. У подкласса Diplopoda, где сегменты двойные, остий по две пары на один сегмент. Сердце продолжается в головную аорту, направляющуюся к мозгу. У губоногих кровеносная система сложно устроена: аорта отдает от себя на пути к мозгу артериальное кольцо, огибающее кишку и впадающее в брюшной продольный сосуд, лежащий над брюшной нервной цепочкой. От каждой камеры сердца отходят, кроме того, 2 боковые артерии. Сердце подвешено к стенкам тела при помощи особых крыловидных мышц. Вес отходящие от сердца сосуды более или менее богато ветвятся, но затем обрываются, и гемолимфа попадает в лакуны миксоцеля, т. е. в промежутки между органами. Из лакун она поступает в околосердечный участок полости тела и оттуда вновь в сердце. Сердце гонит гемолимфу от заднего конца к переднему, в брюшном сосуде она движется в противоположном направлении.
Половая система. Все многоножки раздельнополы. Половые железы лишь в редких случаях сохраняют (некоторые пауроподы) первоначально парный характер и обычно сливаются в непарное образование различного вида. Так, семенник, например, имеет вид массивного образования с лопастями по краю или длинной тонкой трубки, или состоит из 11—12 пар маленьких долек, соединенных общим половым протоком. Яйцевод и семяпровод у кивсяков (см. рис. 305, Г, Д) в начальной части представляют непарные протоки. Направляясь вперед, они раздваиваются и открываются наружу на брюшной стороне второго (не считая шейного) туловищного сегмента. На этом же сегменте расположено половое отверстие у симфил и пауропод.
У представителей подкласса Chilopoda половой проток в начальной части непарный, может образовывать две ветви, которые затем обязательно сливаются. Половое отверстие расположено на предпоследнем сегменте .туловища (см. рис. 305, Б, В).
С половой системой многоножек связан ряд дополнительных образований. Так, в семяпровод часто впадают длинные мешковидные семенные пузырьки. Женская половая система может быть снабжена семяприемниками. Часто развиваются особые придаточные железы (см. рис. 305, Б, В).
Способы оплодотворения многоножек разнообразны. В более простом случае самец подвешивает на выделенную им паутину капельку семенной жидкости или настоящий сперматофор, которые позднее подбираются самкой. Иногда происходит копуляция, причем семенная жидкость в таком случае вносится в половое отверстие самки конечностями самца (чаще всего для этого служат специализированные конечности — гоно-подии).
Развитие.Яйца многоножек крупны и богаты желтком, соответственно с чем они испытывают частичное, поверхностное дробление. Постэмбриональное развитие Myriapoda может протекать по двум несколько различным типам,
Первый тип, или настоящее прямое развитие, встречается у части представителей подкласса Chilopoda (Geophilus, Scolopendra): молодое животное вылупляется из яйца, обладая полным числом туловищных сегментов и конечностей, т. е. вполне походит на материнский организм. Второй тип, или развитие с анаморфозом, имеется у прочих губоногих и
|
| Рис. 306. Шестиногая личинка кив-сяка lulus (Diplopoda) (no Ферхе[\): / — голова, 2 — тель'сон. 3 — зачатки ног, 4 — ноги, 5— антенна; I—V — сегменты туловища |
двупарноногих. В этом случае животное вылупляется с неполным числом туловищных члеников, восполняемых при ряде линек. С каждой линькой к имеющимся сегментам прибавляются позади последнего сформированного сегмента членики, следующие за ним по порядку. Образование их идет за счет зоны роста, которая лежит непосредственно впереди тельсона (т. е. там же, где и у личинок раков). Молодь анаморфных видов подкл. Chilopoda вылупляется с 12 парами туловищных конечностей, молодь подкл. Diplopoda — всего с 3 передними парами ходных ножек, за которыми следует несколько безногих сегментов. Эта шести-ногая стадия (рис. 306) напоминает личинок многих насекомых, когда они еще лишены зачатков крыльев.
Экология.Многоножки преимущественно ночные, избегающие дневного света животные, прячущиеся под корой, камнями и т. п. Кивсяки очень неповоротливы и медлительны, тогда как губоногие, наоборот, ловки и отличаются быстротой движений.
Многие многоножки обнаруживают заботу о потомстве. Они или откладывают яйца в особые гнездышки, приготовленные из земли или другого материала, или свертываются спирально вокруг отложенной кучки яиц и остаются в таком положении по нескольку недель, не принимая пищи, пока не вылупится молодь.
На севере разнообразие многоножек невелико. На юге — в Крыму, на Кавказе, в Средней Азии количество видов их увеличивается. Самые крупные сколопендры и кивсяки — до 28 см длиной и в палец толщиной— встречаются лишь под тропиками. Наиболее мелкие многоножки имеют всего 1—3 мм в длину. Все многоножки, кроме подкл. Chilopoda, совершенно безобидны. Укусы крупных губоногих, например Scolopendra, могут быть болезненными.
Классификация.Многоножки делятся на 4 подкласса (иногда им придают значение самостоятельных классов): симфилы (Symphyla), пауро-поды (Pauropoda), двупарноногие, или кивсяки (Diplopoda), и губоногие (Chilopoda).
ПОДКЛАСС I. СИМФИЛЫ (SYMPHYLA)
Около 150 видов. Мелкие животные с простыми усиками и тремя парами ротовых конечностей. Глаза отсутствуют. Стигмы в числе одной пары располагаются на голове. Конечности имеются не на всех сегментах.
Типичный представитель этой группы Scolopendrella irnmaculata — маленькая (8 мм) многоножка, встречающаяся в лесной полосе Средней и Южной Европы (см. рис. 301, А).
ПОДКЛАСС II. ПАУРОПОДЫ (PAUROPODA)
Около 350 видов. Мелкие формы с антеннами, трехраздельными на конце, и двумя парами ротовых конечностей: мандибул и слившихся в непарную пластинку (гнатохи-лярий) первой парой челюстей. За головой следует безногий «шейный сегмент», соответствующий сегменту третьей пары ротовых конечностей; 10 туловищных сегментов, последний туловищный сегмент безногий. Представитель: Pauropus silvaticus (см. рис. 301, Д) длиной 1 мм. Широко распространены в Европе.
ПОДКЛАСС III. ДВУПАРНОНОГИЕ, ИЛИ КИВСЯКИ (DIPLOPODA)
Около 8000 видов. Тело часто цилиндрическое, усики простые короткие. Ротовых конечностей две пары: мандибулы и гнатохилярий. «Шейный сегмент» лишен конечностей: следующие за ним три первых туловищных сегмента несут по одной паре ножек. Все прочие, за исключением 1—3 последних безногих сегментов, обладают двумя парами конечностей. Представители: Schizophyllum sabulosum (см. рис. 304), распространен по всей Европе; Polydesmus (см. рис. 301, Г) и др.
ПОДКЛАСС IV. ГУБОНОГИЕ (CHILOPODA)
Около 2800 видов. Тело плоское, усики простые, большей частью длинные. Ротовых конечностей три пары: мандибулы и две пары челюстей. Первая пара туловищных конечностей превращена в ядовитые ногочелюсти. Туловищные сегменты несут но одной паре ножек.
Обычным представителем губоногих служит костянка (Lithobius forficatus; см. рис. 301, В) — многоножка, встречающаяся всюду — под корой, под камнями; длина 2—2,5 см; состоит из 19 сегментов (не считая головных). Другая, гораздо длиннее, желтого цвета многоножка Geophilus нередко попадается песной, особенно на огородах и т. п., нападает не только на насекомых, но и на небольших дождевых червей. У нас на юге встречается довольно крупная Scolopendra marsitcins (до 12 см), укусы которой очень болезненны для человека.
В ископаемом состоянии известны преимущественно представители подкл. Diplo-poda; например Archidesmus из силурийских, Euphoberia из каменноугольных отложений и др. Древнейшие губоногие из сем. Euscolopendridac обнаружены в всрхнекамен-ноугольных слоях.
КЛАСС II. НАСЕКОМЫЕ (INSECTA)
Класс насекомых включает всех трахейнодышащих членистоногих, обладающих тремя парами ног. Тело их отчетливо подразделяется на голову, грудь и брюшко. Большинству представителей класса присуща способность к полету — единственная в этом смысле группа среди всех беспозвоночных.
Класс насекомых богаче всех прочих классов по числу представителей. На долю Insccta приходится около 70% общего числа известных видов животных. По разным источникам число изученных видов насекомых колеблется от 600000 до 1 500000. Тем не менее и до сих пор в научной литературе ежегодно описываются новые формы. Если принять во внимание, что многие виды насекомых встречаются в громадном количестве особей, то становится понятным, какую огромную роль насекомые играют в наземных биоценозах. Естественно, что и практическое значение насекомых чрезвычайно велико.
Строение и физиология.Тело взрослых Insecta разделено на голову, грудь и брюшко (рис. 307). Сегменты головы слиты в общую массу, тогда как членики груди и брюшка бывают более или менее хорошо различимы. Голова состоит из акрона и 4 сегментов, грудь всегда из 3, брюшко же в своем наиболее полном составе содержит 11 сегментов и тельсон. Голова и грудь несут конечности, брюшко иногда сохраняет лишь их рудименты.
Голова одета общей хитиновой капсулой, резкий пережим, или шейка, отделяет ее от груди, с которой она большей частью соединена под-
|
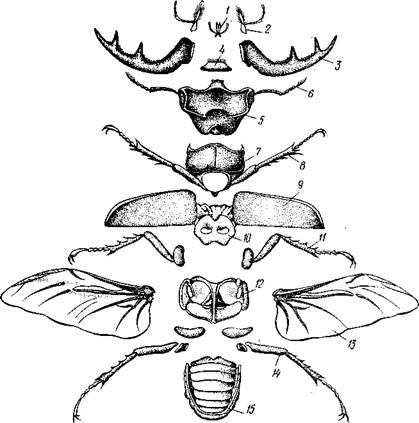
Рис. 307. Расчленение жука-оленя Lucanus cervus (из Гилярова):
/ — нижняя губа, 2 — нижняя челюсть. 3 — мандибула. 4 — верхняя губа, 5 — голова, 6 — сяжки, 7 — передпегрудь, 8 — первая пара грудных конечностей, 9 — передняя пара крыльев — надкрылья, 10 — среднегрудь, // — вторая пара грудных конечностей, 12 — заднегрудь, 13 — задние крылья, 14 — третья пара грудных конечностей, 15 —
брюшко
|
|
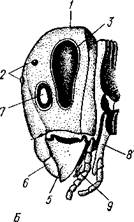
Рис. 308. Голова насекомого спереди (Л) и сбоку (Б) (из Кенигсмена): / — головная капсула, 2 — простые глазки, 3 — фасетированный глаз, 4 — антенна (сяжки), 5 — мандибула, 6 — верхняя губа, 7 — место прикрепления сяжка, S — : нижняя губа, * — нижняя челюсть
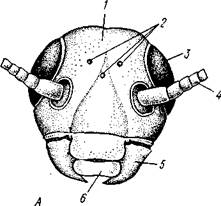
вижно. На нижней поверхности головы или на ее переднем конце помещается рот. По бокам головы расположены два больших сложных глаза, между которыми могут находиться несколько мелких одиночных глазков (рис. 308).
|
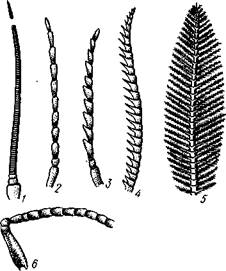
Голова несет 4 пары придатков. От верхней стороны ее отходит пара антенн — придатков акрона. Они гомологичны таковым многоножек, но более разнообразны по форме. Их также часто называют «усиками», или «сяжками». Сяжки всегда состоят из одного ряда члеников. Различают щетинковидпыс сяжки, нитевидные, пильчатые, гребенчатые, перистые, коленчатые и др. (рис. 309). Усики снабжены многочисленными рецепторами, воспринимающими различные типы раздражений. Главным образом это органы осязания и обоняния.
| Рис. 309. Сяжки насекомых (из Кс-нигсмена): / — щетинковидный, 2 — нитевидный, 3 — пильчатый, 4 — гребенчатый, 5 — перистый, 6 — коленчатый |
Интеркалярный сегмент (I сегмент тела) не только не обладает никакими придатками, но и подвергается почти полной редукции. Прочие три пары придатков — конечности II, III и IV сегментов, вошедших в состав головы. Конечности располагаются вокруг ротового отверстия и в совокупности с рядом других образований, имеющих независимое от конечностей происхождение, формируют довольно сложно устроенные у насекомых ротовые аппараты; последние представляют единое функциональное целое.
В соответствии с крайним разнообразием способов питания ротовые аппараты многих групп насекомых отличаются по строению. Однако изучение морфологии и принципов действия этих образований показывает, что многообразные ротовые части Insecta — результат изменений одного общего основного типа. Последний, в свою очередь, возник путем специализации обычных конечностей членистоногих. Захват, удержание и измельчение пищи становятся их главной задачей.
Решая вопрос об исходном у насекомых типе ротовых аппаратов, естественно исходить как из строения, так и из функциональных особенностей последних. Общность плана строения ротовых конечностей и ход-ной ноги, легкость установления гомологии между их отдельными частями— показатели примитивности ротового аппарата в целом. О многом может сказать и характер пищи.
По-видимому, первичным для насекомых было питание более или менее твердым органическим детритом растительного или животного происхождения, что определило появление хищничества или настоящей растительноядности. Но во всех этих случаях животному необходимо захватывать и разгрызать пищевой объект. Поглощение жидкой пищи (кровь животных, соки и нектар растений, жидкие разлагающиеся органические вещества и т. п.) обусловило появление принципиально иных приспособлений.
|
Этим условиям полностью отвечают встречающиеся у прямокрылых, тараканов, жуков и т. п. ротовые аппараты грызущего типа. Их первичный характер доказывается наиболее богатой расчлененностью отдельных конечностей, сходством с ротовыми органами многоножек и тем, что они встречаются у личинок многих отрядов насекомых (например, чешуекрылых) , у которых во взрослом состоянии ротовые аппараты других типов.
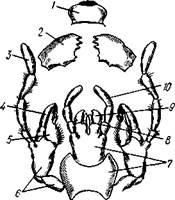
| Рис. 310. Ротовой аппарат грызущего типа черного таракана (по Гертвигу): I — верхняя губа, 2—верхняя челюсть—мандибула, .3—нижнечелюстной щупик, 4 — наружная лопасть нижней челюсти, 5 — внутренняя лопасть нижней челюсти, 6 — нижняя челюсть, состоящая из стволика и основного членика, 7 — нижняя губа, состоящая из подбородка и подподбородка, 8 — внутренняя лопасть нижней губы, 9 —• |
В состав ротового аппарата грызущего типа (рис. 310) входят следующие образования. Спереди рот прикрыт «верхней губой», представляющей складку покровов головы и образующей переднюю стенку ротовой полости. Верхняя губа закладывается независимо от конечностей и не гомологична им. За ней следует пара верхних челюстей— жвал, или мандибул. Это две толстые нерасчлененные пластинки, зазубренные по внутреннему краю и играющие наиболее важную роль в размельчении пищи. Мандибулы являются конечностями II сегмента головы. Конечности III и IV сегментов — максиллы, или нижние челюсти. Они члени-сты н этим напоминают О происхождении ОТ ходных ног. Первая пара нижних челюстей
г г
раСПОЛОЖеНЗ ПО бОКЗМ рТЗ, ПрИЧбМ КЭЖДЗЯ челюсть СОСТОИТ ИЗ ДВуЧЛбНИКОВОГО ОСНОВа-
ния и сидящих на его вершине трех придатков: двух лопастей, внутренней и наруж-
Дата добавления: 2015-01-24; просмотров: 2317;