Целом, 2 — половая воронка. 3 — протонефридий, 4 — метанефридий
В тех случаях, когда протонефридии уже превратились в метанефри-дии, дело может обстоять двояко: иногда половые протоки и метанеф-ридии остаются обособленными; у части же полихет происходит слияние половой воронки с внутренним (отверстым) концом метанефридия. В результате получаются тоже нефромиксии, но несколько иного свойства, возникающие из слияния половых воронок не с протонефридиями, а с метанефридиями (рис. 219, Б; 220, Г). Нередко, особенно у сидячих Polychaeta, число выделительных каналов сокращается; они сохраняются всего в нескольких сегментах. Обычно, у сидячих полихет происходит физиологическая дифференциация нефромиксисв на чисто выделительные и служащие для выведения половых продуктов.
Кроме нефридисв есть другие образования, играющие выделительную роль. В некоторых местах, главным образом на стенках кровеносных сосудов, перитонеальный эпителий состоит из крупных клеток, содержащих множество желтых зерен, — это накопляющиеся в клетках нерастворимые продукты обмена веществ (гуанин или соли мочевой кислоты). Наполнившись экскретами, эти, как их называют, хлорагогенные клетки отмирают, а содержимое их поступает в целом и оттуда через нефридии — наружу.
Нервная система обнаруживает ряд ступеней усложнения. Типичная центральная нервная система (см. рис. 217, А) состоит из парных мозговых ганглиев, отходящих от них и огибающих глотку двух окологлоточных коннективов и парного брюшного нервного ствола. Коннективами называются нервные стволы, соединяющие разноименные ганглии (надглоточные и подглоточные или ганглии двух соседних сегментов). Поперечные нервные стволы, связывающие ганглии одного сегмента, называются комиссурами.
У примитивных форм обе половины брюшного ствола еще широко расставлены и имеют равномерную обкладку из нервных клеток. Далее, у части Polychaeta правый и левый брюшные стволы начинают сближаться к срединной линии; в каждом сегменте на них появляется по скоплению ганглиозных клеток, и оба ганглия сообщаются поперечной перемычкой (комиссурой) —получается брюшная нервная система, метко названная нервной лестницей (см. рис. 212). У большинства полихет стволы сближаются еще больше, оба ганглия каждого сегмента сливаются, сами же стволы между ганглиями либо еще остаются разделенными, либо тоже сливаются, и тогда первичную парность брюшного ствола можно определить только на поперечных разрезах. Нервная система такого устройства называется брюшной нервной цепочкой.
Другое постепенное усложнение нервной системы заключается в ее погружении из кожного эпителия в полость тела. Во время эмбрионального развития нервная система закладывается в виде утолщения эктодермы. У некоторых Polychaeta брюшная нервная система и во взрослом состоянии остается в кожном эпителии, не меняя своего положения. У многих полихет нервные стволы уходят под кожу и даже под кожно-мускульный мешок, располагаясь в полости тела. В некоторых случаях, когда ряд сегментов сливается друг с другом (например, в области глотки), может происходить и соответственная концентрация нервных узлов.
От центральной нервной системы отходят многочисленные нервы. Головной мозг посылает от себя нервы к антеннам, пальпам и глазам, а каждый ганглий брюшной нервной цепочки иннервирует различные органы соответствующего сегмента.
Органы чувств лучше всего развиты у бродячих полихет. Кроме эпителиальных чувстительных клеток, рассеянных в коже, есть специальные
9* 259
органы осязания и химического чувства. Таковыми служат антенны, пальпы, ресничные ямки, располагающиеся на простомиуме, и чувствительные усики параподий.
Органы чувства равновесия редки, однако у некоторых полихет, особенно у сидячих, в передних сегментах тела имеется от 1 до 5 и более пар статоцистов.
Органы зрения имеются почти у всех многощетинковых. Чаще всего глаза расположены на спинной стороне простомиума в числе 2 или 4 и
|
| 8 |
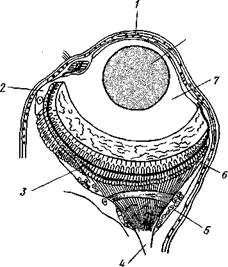
| Рис. 221. Глаз полихеты Alciope (из Ливанова): / — роговица. 2— клетка, выделяющая стекловидное тело, 3 — светочувствительные палочки, 4 — зрительный нерв, 5 — зрительный ганглий, 6 ~ сетчатка, 7 — стекловидное тело, S — хрусталик |
иннервируются от средней части головного мозга. Эти надмозговые глаза в наиболее простом случае представляют бокаловидное впячи-вание эктодермы с суженным отверстием. Выстилающий такую глазную ямку эпителий играет роль сетчатки, а по краям впячивания переходит в обыкновенный кожный эпителий. Сетчатка слагается из клеток двух сортов. Одни из них светочувствительны, и они-то, собственно, и являются настоящими рети-нальными клетками. Каждая такая клетка на конце, обращенном в полость впячивания, т. е. к источнику света, несет светочувствительную палочку, а на другом конце продолжается в нервное волокно. Нередко, особенно у планктонных форм, например у Alciope (рис. 221), глаз сильно усложняется: обособляется от кожи, превращаясь в замкнутый глазной пузырь, а внутри него дифференцируется, кроме двояковыпуклого хрусталика, еще прозрачное стекловидное тело. Все описанные глаза необращенного, или неинвертированного, типа. Но помимо надмозговых глаз или (при их редукции) вместо них у полихет нередко развиваются глаза на других частях тела, притом обладающие иным строением. Так, у многих сидячих форм, живущих в трубках, развиваются многочисленные глаза на пальпах, превращенных в жабры. У некоторых мелких форм (АтрЫсога и др.), ползающих задним концом вперед, развиваются глазки возле порошицы. Эти «вторичные» глаза различного строения.
Половая система устроена крайне просто. Многощетинковые кольчецы раздельнополы, наружных отличий между полами нет. Половые железы формируются во всех (кроме передних и самых задних) или лишь в некоторых плодущих сегментах под слоем перитонеального эпителия. Чаще всего у основания параподий или по соседству с нефридия-ми половые клетки усиленно делятся и образуют на стенке целома местное набухание или половую железу (см. рис. 216; 217, Б). Развивающиеся гонады сначала прикрыты тонким слоем перитонеального эпителия, который впоследствии лопается, и образующиеся половые клетки попадают в целом, где они свободно плавают в полостной жидкости и достигают полной зрелости. У некоторых Polychaeta никаких половых протоков нет, так что живчики или яйца выводятся наружу просто через раз-
|
рыв стенки тела. У очень немногих полихет (сем. Capitellidae) имеются самостоятельные половые воронки с короткими выводными' каналами, открывающимися наружу (см. рис. 220, В). Наконец, в большинстве случаев эти воронки, как сказано, вступают в сообщение с нефридиями, и тогда нефридии несут двоякую функцию — выделение экскретов и выведение наружу половых продуктов. Оплодотворение обыкновенно наружное.
| Рис. 222. Тихоокеанский палоло Eunice viridis (из Фаусека). Передний, более толстый отдел тела, — атокный, задний, более тонкий, — эпитокный |
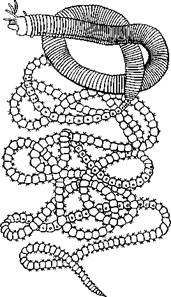
Нередко у полихет в тесной связи с половым размножением находится бесполое, приуроченное к периоду созревания половых продуктов. Переходом к этому размножению можно считать так называемые эпи-токные формы многощетинковых. Обычно наступление половой зрелости не отражается па организации червя. Иногда, однако, плодущие сегменты претерпевают сильную модификацию и в виде так называемой эпигокной части резко отличаются от остального атокного участка животного. Чаще всего эпитокной становится задняя половина червя. Изменения эпитокного участка сказываются в более сильном развитии параподий и щетинок, в сильной рудиментации кишечника, изменении окраски и т. д. Эпитокные половозрелые стадии некоторых представителей семейств Nereidae, Eunicidae и других поднимаются со дна и при помощи мощных веслообразных пара-подий свободно плавают, приступая к половому размножению. Таков знаменитый па-лоло Тихого океана, который периодически в несметном количестве поднимается со дна и роится в поверхностных слоях воды, представляя в это время лакомую пищу для туземцев (Eunice viridis; рис. 222). У других видов задняя половина тела отрывается от передней, регенерирует на месте разрыва новую голову и, обладая более мощными параподиями, уплывает. Бесполая половина остается на прежнем месте и регенерирует задние сегменты.
У Autolytus (рис. 223, A), Myrianida и некоторых др. эпитокная половина может еще до отделения от атокной образовать голову, а бесполая—-начать регенерацию задних сегментов: животное имеет вид двух особей разного строения, соединенных зоной регенерации. Иногда, наконец, еще до отделения первого сформировавшегося полового индивида впереди него за счет зоны регенерации образуется вторая, третья и т. д. до 30 половых особей, расположенных в один ряд (рис. 223, А); получается временная цепочка особей (Autolytus и некоторые другие). Затем половые особи отделяются и уплывают (рис. 223, Б, В).
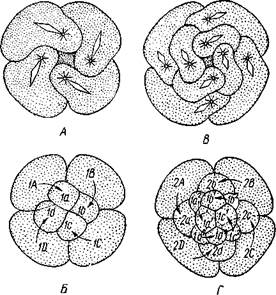
Развитие.Дробление яйца полное. Сначала двумя последовательными меридиональными делениями яйцо рассекается на 4 приблизительно одинаковых бластомера: А, В, С и D, причем В отвечает будущей брюшной, a D — будущей спинной стороне зародыша. Затем эти 4 клетки делятся наклонной бороздой на 4 мелкие верхние (анимальные) и 4 нижние крупные (вегетативные) клетки. Получается, как говорят, квартет микромеров и квартет макромеров (рис. 224). Микромеры, смотря по их происхождению от того или иного макромера, обозначают как la, \b, lc
и Id (рис. 224),макромеры—\А — ID. Дальнейшее дробление состоит в последовательном отделении от макромеров по направлению к ани-мальному полюсу вслед за первым еще 2-го, 3-го и 4-го квартетов микромеров, при отделении каждого нового квартета клетки ранее образовавшихся квартетов тоже делятся.

|
Рис. 223. Размножение полихет. А — бесполое размножение по-лихеты Autolytus (по Грассе); 5 и В — получающиеся посредством концевого почкования половые особи (5 — самка и В — самец) тех же червей (по Свешникову)
Важная особенность отделения микромеров заключается в изменении направления веретена деления: анимальный конец его отклоняется то по ходу часовой стрелки, то против нее, если наблюдение вести с ани-мального полюса яйца. Если при образовании 1-го квартета ядерные веретена в бластомерах наклонены в одну сторону, то при отделении 2-го квартета все они оказываются отклоненными в другую, и т. д. Вследствие этого верхние продукты деления ложатся каждый раз не прямо над нижними, а несколько чередуясь с ними, как бы в шахматном порядке. Например, микромер \а лежит не целиком над макромером 1Л, но отчасти налегает и на соседний макромер 1В. Такое дробление называется спиральным, так как если во время деления микромера мысленно продолжить ось ядерного веретена по сферической поверхности яйца, то получится спиральная линия.
|
| Рис. 224. Схема спирального дробления. Вид с верхнего полюса зародыша. А — переход от четырех- к восьмиклеточной стадии; Б — стадия восьми клеток; В — переход к стадии 16 клеток; Г — зародыш на стадии 16 клеток (из Зивинга) |
Уже на ранних стадиях дальнейшее развитие каждого эластомера точно определено. В общем, первые 3 квартета микромеров идут на образование эктодермы личинки, один из микромеров 4-го квартета — потомок клетки D, обозначаемый 4d, производит целоми-ческую мезодерму, а остальные микромеры 4-го квартета и все макромеры дают энтодерму. Ввиду такого раннего определения дальнейшего развития бластомеров дробление кольчецов называется детерминативным. В результате дробления получается шаровидная бластула, из которой посредством погружения внутрь бла-стоцеля группы крупных эн-тодермальных клеток происходит гаструла. Бласто-пор гаструлы помещается на ее вегетативном полюсе. Затем бластопор вытягивается по одной из сторон гаструлы (будущей брюшной) к ее экватору, а сзади замыкается. Его передняя часть превращается в ротовое отверстие. На нижнем полюсе зародыша образует-
ся порошица. Постепенно зародыш превращается в характерную для полихет личинку — трохофору.
Трохофора (рис. 225, А)—типичная планктонная личинка, плавающая при помощи ресничек. Тело ее имеет более или менее шаровидную или эллиптическую форму. На переднем (анимальном) полюсе личинки развивается чувствительный теменной султан длинных ресниц, сидящих на группе эктодермальных клеток — теменной пластинке. По экватору личинки впереди рта расположен характерный предротовой венчик ресничек— прототрох. Иногда позади рта развит менее мощный послеро-товой венчик. Кишечник начинается ртом посредине брюшной стороны личинки и заканчивается порошицей на ее заднем полюсе и состоит из трех отделов, причем передняя и задняя кишка образуются впячиванием эктодермы, а средняя кишка формируется из энтодермы. Между кишечником и стенкой тела находится первичная полость тела, пересекаемая тонкими мышечными волокнами. По бокам кишечника лежит пара маленьких протонефридиев.
Мезодермальные органы личинки (главным образом мускульные волокна) развиваются из нескольких клеток, лежащих у краев бластопора и называемых мезенхимой. Другой мезодермальный зачаток представлен двумя крупными мезодермальными клетками — первичными мезоб-ластами (иногда их называют телобластами)—потомками бластомера 4d, лежащими по бокам от кишечника.
После некоторого периода планктонной жизни трохофоры начинается ее метаморфоз. Заднее (вегетативное) полушарие личинки значитель-
но вырастает в длину и подразделяется сразу на несколько (3, 7, 9—13) сегментов. На сегментах развиваются параподии и щетинки или появляются ресничные пояски. К этому времени обе первичные мезодермальные клетки, усиленно размножаясь, дают два лежащих по бокам от кишечника клеточных тяжа — мезодермальные полоски (рис. 225, Б). Вско-
|
| 13 |
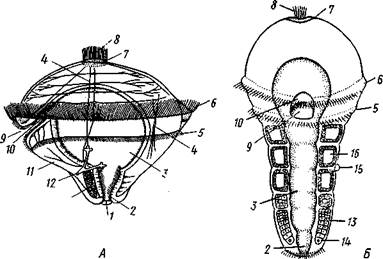
Рис. 225. Развитие Poltjgordius. A — трохофора, вид сбоку (по Гатчеку); Б — метаморфоз трохофоры (по Вурмбаху):
1 —анус, 2 — задняя кишка, 3 — средняя кишка, 4 — мышцы, 5 — послеротовой и 6 — предротовой венчик ресничек (нрототрох), 7 — теменная пластинка, 8 — теменной султан, 9 — рот, 10—передняя кишка, 11— нижнее полушарие трохофоры, 12—протонефридий, 13 —мезодермальная полоска, 14 — мезобласт, 15 — диссепи-
мент, 16 — целом
ре под влиянием наружной сегментации мезодермальные полоски расчленяются на парные группы клеток, так что в каждом сегменте оказывается своя пара мезодермальных клеточных зачатков. Последние сначала компактны, затем в них появляется полость — зачаток вторичной полости тела, а ограничивающая ее клеточная стенка есть стенка цело-мического мешка. В каждом сегменте, таким образом, развивается пара целомических мешков. В процессе метаморфоза часть клеток теменной пластинки трохофоры погружается под покровы и образует головной мозг. На брюшной стороне в виде парного валика эктодермы закладываются брюшные нервные стволы. В дальнейшем они вступают в связь с головным мозгом при помощи окологлоточных коннективов. Из эктодермы развиваются и органы чувств — глаза, пальпы.
Так, из несегментированной, первичнополостной трохофоры формируется следующая личиночная стадия — метатрохофора (рис. 226), характеризующаяся сегментацией и метамерным целомом. Тело сформированной метатрохофоры состоит из головной лопасти (простомиума), нескольких сегментов и маленькой анальной лопасти (пигидиума) с порошицей на конце. Головная лопасть представляет собой почти неизменившееся переднее предротовое полушарие трохофоры, а анальная лопасть— самый нижний участок трохофоры, лежащий позади уровня первичных мезодермальных клеток. Таким образом, оба отдела метатрохофоры по природе резко отличны от сегментов туловища. Существенная
особенность сегментов метатрохофоры заключается в том, что все они возникают одновременно.
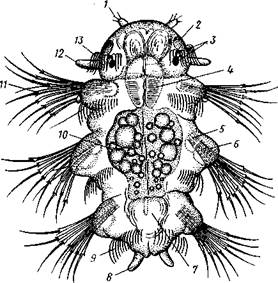
|
| Рис. 226. Метатрохофора Nereis pelagica (no Рунштрому): / — щупальце. 2 — пигментная клетка. 3—глаза, 4 — глотка, 5 — щетинконосный мешочек параподии, 6 — па-раподия, 7 — задняя кишка, 8 — анальный усик, 9 — ресничный поясок, 10 — средняя кишка, 11 —щетинки, 12 — пальп, 13 — прототрох |
|

|
|
|
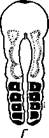
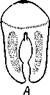
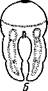
| Рис. 227. Схема соотношений между мезодермой лар-вальных и постларвальных сегментов при развитии Polychaeta (по П. Иванову). А — превращение тро-хофоры в мстатрохофору, мезодермальные полоски не сегментированы; Б — Метатрохофора с тремя лар-вальными сегментами, мезодермальные полоски сегментированы; В — появление на заднем конце тела эктодермальной зоны роста; Г — образование постларвальных сегментов из зоны роста (зона роста я целомические мешки постларвальных сегментов зачернены) |
Метатрохофора некоторое время плавает или ведет дойный образ жизни, не меняясь существенно, а затем испытывает дальнейший метаморфоз. На переднем крае анальной лопасти образуется зона роста, клетки которой непрерывно размножаются. Область, лежащая впереди нее, состоит из быстро растущих, еще не дифференцированных тканей. В этой зоне формируются новые сегменты и последовательно один за другим отделяются по направлению кпереди (рис. 227). Процесс продолжается до тех пор, пока не образуется столько сегментов, сколько их у взрослого червя. Иногда первичные мезодермальные клетки сохраняются в зоне роста и дают начало мезодермальным полоскам. Чаше, однако, эти клетки целиком расходуются на образование ие-ломической мезодермы метатрохофоры, и мезодермальные полоски образуются за счет размножения эктодермальных клеток зоны роста. От ме-зодермальных полосок последовательно отделяются парные зачатки цело-мических мешков (рис. 227). Каждый вновь образующийся сегмент получает свою пару таких зачатков, из которых развиваются его целомические мешки, разрастающиеся по бокам кишечника кверху и книзу, постепенно вытесняя собой первичную полость тела. Наконец, правый и левый мешки в каждом сегменте встречаются над и под кишкой по спинной и брюшной линиям, а следующие друг за другом пары мешков тоже соприкасаются своими передними и задними стенками
(рис. 228). Наружным кожно-мускульным листком мешки подстилают эктодерму, а внутренним граничат с кишечником. В результате описанного процесса первичная полость тела личинки замещается целомом. За счет соприкасающихся стенок целомических мешков над и под кишкой образуется спинная и брюшная брыжейки, а на месте соприкосно-
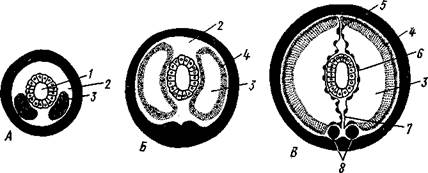
Рис. 228. Развитие целома у кольчатых червей. А—В — поперечные разрезы
трех последовательных стадий развития сегмента (из Матвеева):
/ — кишка. 2 — первичная полость тела. 3 — целом, 4 — наружная стенка целомиче-
ского мешка, 5 — спинной мезентерий, 6 — внутренняя стенка целомического мешка,
Дата добавления: 2015-01-24; просмотров: 2120;