Кишка, 7—половая железа, 8 — анальный мешок, 9 — анальное отверстие
Тело одето однослойным эпителием, выделяющим из своей поверхности кутикулу. Под кутикулой расположен кожно-мускульный мешок. Пе-ритонеальный эпителий ограничивает обширную сплошную полость тела (целом).
Пищеварительная система образует длинный извитой канал, состоящий из передней, средней и задней кишки, открывающийся на заднем конце анусом (рис. 246). Благодаря извилистому ходу длина кишечника
иногда в 10 раз превышает длину тела. В заднюю кишку впадает пара ее выпячиваний: анальные мешки. Они усажены 12—300 мелкими мерцательными воронками, которые открываются в целом, а другим концом в полость анального мешка. Мешки служат для дыхания и для выделения.
Кровеносная система состоит из проходящего над передней частью кишки спинного продольного сосуда, продолжающегося и в хоботок. Этот сосуд впереди раздваивается и дает два боковых хоботковых сосуда. Последние при выходе из хоботка сливаются в общий продольный брюшной сосуд, тянущийся под кишкой до заднего конца тела. Задний конец спинного сосуда сообщается с брюшным при помощи двух попе-
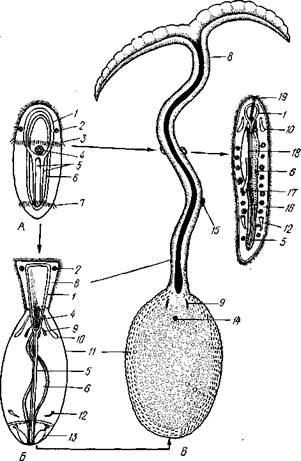
Рис. 247. Фенотипическое определение пола у Bonel-lia viridis. A — плавающая личинка; Б — женская личинка старшего возраста; В — взрослая самка; Г —
самец из матки самки (по Бальтцеру): 1 — окологлоточное нервное кольцо, 2 — глазные пятна, 3 —• прототрох. 4 — передняя кишка, 5 — брюшные нервные стволы, в — средняя кишка. 7 — анальный венчик ресниц, 8 — хоботок, 9 — щетинки, 10 — протонефридий. 11 — тело, 12 — метанефридий, 13—анальный мешок с воронкой. 14 — женское половое отверстие, 15 — личинки на хоботке самки. /6 — целом с развивающимися семенными клетками. 17 — мужская половая воронка, 18 — семенной проток, 19 — мужское половое отверстие
речных сосудов, охватывающих кишечник. Кровь бесцветна и содержит бесцветные лейкоциты.
Нервная система развита довольно слабо. Центральная нервная система состоит из брюшного ствола (см. рис. 246), который на переднем конце туловища раздваивается, огибает кишку и образует окологлоточное нервное кольцо. Итак, общий план строения нервной системы напоминает Polychaeta, no нервный ствол Echiurida на всем протяжении усеян нервными клетками и даже окологлоточное кольцо лишено ганг-лиозных утолщений. Органы чувств, кроме отдельных чувствительных клеток и сосочков в коже, отсутствуют.
Выделительная система. Нефридии представлены троякого рода органами. На стадии личинки трохофоры эхиуриды обладают парой прото-нефридиев, которые во время метаморфоза редуцируются (как и у кл. Polychaeta). У взрослых животных для выделения служат прежде всего анальные мешки с их вороночками, рассматриваемые некоторыми зоологами как метанефридии. Кроме мешков, у взрослых эхиурид имеются более или менее типичные нсфромиксии, открытые в целом мерцательными воронками. Однако главное назначение их-—выведение половых продуктов. Нсфромиксии лежат в передней части тела и открываются наружу на брюшной стороне. Число их варьирует от 1 до 4 пар, но у некоторых форм их значительно больше. Так, например, у крупной Ikeda tenioides с каждой стороны брюшного нервного ствола располагается от 100 до 200 нефридиев. У громадного большинства родов происходит постепенная олигомеризация числа выделительных органов от 4 пар у Thalassema до 1 у самки ВопеШа.
Половая система. Эхиуриды раздельнополы. Гонада непарная, лежит на брюшной стороне в задней трети тела. У родов ВопеШа и Hamingia наблюдается крайне резкий половой диморфизм (рис. 247). Самки ВопеШа— крупные (до 10—15 см) черви зеленого цвета с длинным, раздвоенным на конце хоботком. Самцы ВопеШа были открыты О. Ковалевским. Это маленькие (1—3 мм) червячки, которые часто попадаются
|
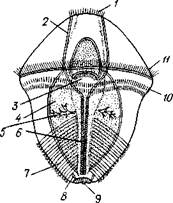
| Рис. 248. Трохофора Ecfiiurus с брюшной стороны (по Гатче- ку): / — теменная пластинка, 2 — глоточный коннсктив. 3 — рот. 4 — кишка, 5—протонефридий, 6 — брюшной нервный ствол, 7—наружная складчатость покровов, 8 — зачаток анального мешка, 9—анус, 10 — послкротовой венчик ресничек, 11 — прототрох |
в полости нефридиев самки и прежде считались ее паразитами. Самцы сплошь покрыты ресничками, напоминая этим турбел-лярий, и по сравнению с самкой сильно упрощены. Ни рта, ни порошицы у них нет, так что кишечник их замкнут слепо; кровеносной системы нет. Для вывода живчиков служит особый семенной мешок, происходящий из средней кишки личинки и нефриди-альной воронки. Анальных мешков нет. Образ жизни самцов оригинален. Сначала они ползают по поверхности хоботка самки; позднее проникают в переднюю кишку, потом в нефридии самки и живут там подолгу в ожидании прохождения через нефридии созревших яиц, которые оплодотворяют, выпуская живчики.
Развитие эхиурид сначала напоминает таковое полихет. Спиральное и детермина-тивное дробление приводит к образованию более или менее типичной трохофоры (рис. 248). Рост трохофоры сопровождается образованием двух мезодермальных полосок. Однако последние не сегментируются, а,
сильно разрастаясь и сливаясь воедино, образуют общий целом. Тело личинки позади прототроха равномерно растет в длину. Личинка свободно плавает в планктоне и лишь после метаморфоза оседает на дно.
Развитие Bonellia.Эксперименты обнаруживают очень характерное влияние условий выращивания личинок Bonellia на определение пола развивающихся из них особей. При воспитывании личинок Bonellia поодиночке из них всегда получаются самки. Если же личинки выращиваются в сосуде с взрослой самкой, то они оседают на хоботок последней, испытывают регрессивный метаморфоз и превращаются в паразитических самцов. Таким образом, Bonellia представляет один из редких примеров животных, у которых пол определяется в метагамнын период, т. е. после оплодотворения половой клетки. Определяющее влияние на развитие личинок у самцов оказывают, по-видимому, выделения покровов хоботка самки, в которых заключается особый зеленый пигмент. Можно изготовить из тканей самки Bonellia экстракт, растворы которого в морской воде действуют на личинок таким же определяющим способом, как и присутствие живой самки.
КЛАСС СИПУНКУЛИДЫ (SIPUNCULIDA)
Сипункулиды — небольшая группа (250 видов) морских червеобразных животных, ведущих главным образом роющий образ жизни или прячущихся в пустых трубках и раковинах других животных. Сипункулиды прежде рассматривались как потомки аннелид, утратившие сегментацию под влиянием роющего образа жизни. В действительности, однако, ни в строении взрослых сипункулид, ни в развитии их нет никаких следов метамерии. Подобно эхиуридам, это первично несегментированные целомические животные (Coclomata).

|
Основные черты организации сипункулид сводятся к следующему: несегментированное тело их делится на узкий, способный вворачиваться и выворачиваться хоботок и более широкое туловище. На вершине хоботка лежит ротовое отверстие, обычно окруженное венчиком коротких щупалец, анус расположен на спинной стороне тела, несколько позади рта. Имеется хорошо развитый кожно-мускульный мешок. Вторичная полость тела имеет вид цельного несегментированного туловищного целома и несообщающегося с ним переднего целоми-ческого кольца, от которого отходят целомические каналы щупалец.
Кровеносная система отсутствует. Органами выделения служат нефромиксии. Сипункулиды раздельнополы. Яйца дробятся по спиральному типу. Из яйца выходит типичная личинка трохофора.
Сипункулиды обитают во всех морях, будучи характерными представителями донной фауны. Их средние размеры составляют несколько сантиметров, самой крупной формой является Siphonomecus multi-cinctus, достигающий 51 см в длину.
Строение.На переднем конце внутри венчика не
больших ресничных щупалец расположено ротовое
отверстие. Передний участок тела вместе со ртом мо
жет вворачиваться внутрь тела при помощи четырех рис 24g Phasco[0^
специальных мускулов-ретракторов. Выворачивание soma" margaritacea
переднего участка тела (хоботка) совершается при (по Гилю):
посредстве давления целомической жидкости и со- / — хобот, г — аналь-
кращения кольцевых мышц стенки тела. Анальное н°\енТиТРщупале11 ~"
отверстие помещается на спинной стороне тела позади основания вворачивающегося хоботка (рис. 249).
Стенка тела. Под довольно плотной кутикулой лежит кожный однослойный эпителий с крупными многоклеточными железами. Кожно-мус-кульный мешок сплошной и состоит из наружного слоя кольцевых, среднего — косых и внутреннего — продольных мышц. Обширная вторичная полость тела выстлана перитонеальным эпителием.
Кишечник состоит из эктодермальной глотки, длинной средней кишки энтодермальной природы и эктодермальной задней кишки. Общая длина кишечника в два раза превышает длину тела. Кишечник направляется назад, делает многочисленные волнообразные изгибы, достигает заднего конца тела, затем поворачивает обратно и таким же извилистым путем идет вперед; задняя кишка заканчивается порошицей недалеко от переднего полюса животного, несколько впереди задних концов мышц-ретракторов (рис. 250). На своем пути кишечник обвивается вокруг особой поддерживающей его связки, которая одним концом прикреплена к стенке тела в области порошицы, другим же — в области заднего полюса животного. Сипункулиды заглатывают морской песок или ил, откуда ими извлекаются органические остатки. Они поглощают большое количество грунта, масса которого может составлять свыше половины общей
массы тела.
|
Нервная система состоит из парного надглоточного ганглия, или головного мозга, из двух окологлоточных коннективов и из непарного, лишенного ганглиозных вздутий брюшного нервного ствола; последний лежит на внутренней стороне стенки тела, вдаваясь в целом. Единственными органами чувств служат околоротовые щупальца.
Дыхание совершается отчасти через поверхность тела, а также через стенки щупалец, которые нежнее остальных покровов. Функции кровеносной системы выполняют лакуны и синусы, имеющиеся в стенке кишечника. Жидкость лакун бесцветна, со-
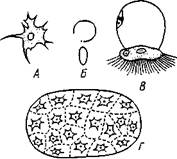
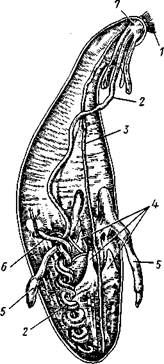
Рис. 250. Анатомия Phascoloso-
tna (из Холодковского): 1 — щупальца, кзади от них торчат перерезанные мускулы-ретрак-торы, 1—кишечник, 3 — брюшной нервный тяж, 4 — перерезанные мускулы-ретракторы переднего конца тела, 5 —нефридии. 6 — задняя кишка. 7 — надглоточный ганглий
Рис. 251. Элементы крови Sipunculus. A — лейкоцит; Б — красное кропя-ное тельце с плоской стороны и в профиль; В — двуклеточная урночка; Г — загадочный диск (по Догелю)
держит лишь амебоидные клетки и не несет дыхательной функции крови. Таковую у сипункулид выполняет жидкость вторичной полости тела — целома.
Целом разделен поперечной перегородкой на два участка. Передний участок очень мал; он охватывает в виде узкого кольца передний конец кишки и посылает от себя выпячивания вперед, внутрь щупалец, и два довольно длинных слепых выроста назад; эти выросты прилегают к кишечнику со спинной и брюшной сторон и раньше ошибочно принимались за кровеносные сосуды.
Жидкость целома розового цвета, мутная от многочисленных клеточных включений (рис. 251). Разнообразие плавающих в ней клеток чрезвычайно велико. Целомическая жидкость содержит прежде всего амебоидные клетки с псевдоподиями. Далее, в ней имеются округлые сплющенные клетки розоватого цвета. Окраска зависит от присутствия в них железосодержащего вещества, гемэритрина, близкого к гемоглобину. Таким образом, эти клетки можно приравнять к красным кровяным тельцам позвоночных, а сама полостная жидкость, несомненно, играет роль крови.
Особенно любопытными включениями целома являются так называемые урночки. В наиболее простом случае — это двуклеточные тельца, одна клетка которых имеет вид прозрачного купола, а другая — форму прижатой к основанию купола вогнутой лепешки, усаженной многочисленными ресничками. При помощи ресничек урночки быстро плавают в полостной жидкости. У некоторых сипункулид имеются более крупные многоклеточные урночки. Урночки формируются за счет стенок целома, его перитонеума. Сначала они сидят на стенках полости тела на ножке, а потом отрываются и уже свободно плавают в целоме. Урночки принимают участие в выделении экскретов. Их мерцательная клетка выделяет липкое вещество, которым склеиваются накопляющиеся в полости тела в виде желтых зерен продукты обмена веществ, а также нагруженные этими отбросами мелкие амебоидные клетки. Эти кучки отбросов вместе с самими урночками спаиваются в комки более крупного калибра, так называемые желтые тела, которые, по-видимому, выводятся потом через нефридии.
Кроме урночек, в целоме плавают также диски —прозрачные овальные пластинки, состоящие из 2—64 многоугольных клеток. Судя по некоторым наблюдениям, диски служат как бы живыми пластырями, которые прикладываются к местам кишечника, поврежденным острыми обломками содержимого кишечника (песчинками и т. п.). На месте поранения скопляются десятки дисков, которые приклеиваются к стенке кишки при посредстве амебоидных клеток целома (сами по себе диски неподвижны). Диски следует рассматривать как полезное приспособление, вызванное особым характером пищи, очень часто повреждающей кишечник. Наконец, в полостной жидкости сипункулид обыкновенно имеются еще созревающие половые клетки.
Выделительная система представлена нефромиксиями, число которых варьирует от 1 до 3. Они лежат в передней трети тела и открываются наружу несколько впереди порошицы, на брюшной стороне. Нефридии имеют вид крупных темно-коричневых мешков с мерцательной воронкой, открывающейся в полость тела. Помимо выделения, нефридии служат для выведения наружу половых продуктов.
Половая система. Сипункулиды раздельнополы. Половые железы развиваются в виде складчатых валиков из перитонеального эпителия у основания брюшных мускулов-ретракторов. Половые клетки еще на ранних стадиях отрываются от половых желез и падают в целом, где
заканчивается их созревание. Оплодотворение происходит в морской воде.
Развитие.В эмбриональном развитии следует отметить спиральное детерминированное дробление, образование личинки трохофоры и формирование двух симметричных мезодермальпых полосок из первичных мезобластов. Полоски не сегментируются, клетки их расходятся, и между ними образуется обширная общая вторичная полость тела. В развитии сипункулид, таким образом, нет никаких следов сегментации, или метамерии. Трохофора сипункулид постепенно превращается во взрослое животное, испытывая разрастание тела преимущественно в своей задней половине, в области, лежащей позади анального отверстия, которое уже у личинки сдвинуто на спинную сторону и вперед.
Экология.Взрослые Sipunculida ведут, как сказано, малоподвижный или скрытый образ жизни. Яйца и личинки их проходят свое развитие в планктоне, что облегчает распространение этих животных морскими течениями. Сипункулиды встречаются почти во всех морях и океанах, за исключением некоторых внутренних морей с более или менее опресненной водой (в Балтийском и Черном морях они отсутствуют).
Представители: Phascotosoma margarilacca — с многими ротовыми щупальцами до 19 см длиной, обитает в иле, Slpunculus nudus — до 25 см длиной, Phascolion slrombi — до 2 см длиной, живет в пустых раковинах лопатоногого моллюска Dentalium, Siphonotnecus mullicinctus — самая крупная сипункулида (до 50 см), встречается в Тихом океане у берегов Калифорнии.
Филогения.Родство сипункулид с кольчатыми червями не подлежит сомнению. О нем свидетельствуют спиральное дробление яйца, наличие трохофоры и телобластический способ развития целома. Однако старые данные о том, что у некоторых сипункулид мезодермальные полоски временно сегментируются, не подтвердились. Вероятно, класс Sipunculida представляет реликтовую группу, близкую к вымершим первичным Protostomia с общим, несегмснтированным целомом, от которых произошли аннелиды и связанные с ними группы (моллюски и членистоногие).
Дата добавления: 2015-01-24; просмотров: 1856;